MICROORGANISM VIRULENCE
Endotoxin
The unique outer membrane of gram-negative bacteria, containing phospholipids, LPS, and proteins, has been implicated as a major player in pathogenesis. The LPS portion of the envelope is both a major cell surface antigen and a major molecule triggering host response to bacterial infection. It is made up of three regions: the O-specific region (region I), responsible for the O or somatic antigenicity of the species and subspecies; the core polysaccharide (region II); and lipid A (region III), which is closest to the cell wall. The term “endotoxin” was used before discovering that it is the lipid A portion of the LPS. Different sugar sequences, linkage groups, and substituents cause different antigenic (serologic) specificities. The presence of O-antigen in Serratia marcescens enhances its adherence to inert and biologic surfaces.44 Region II is less variable than region I. Region III is highly conserved among eukaryotes, with little microheterogeneity among genera and species. Several members of the family Enterobacteriaceae express additional antigenic polysaccharides in the form of capsular or K antigens.
Lipid A is crucial in that it is responsible for the pathophysiologic effects associated with gram-negative bacterial infections and bacteremia, including pyrogenesis and hematologic, immune, and endocrinologic effects. Systemic exposures to endotoxin may result in hypotension, disseminated intravascular coagulation, and death. Most of these effects are mediated by cytokines and some by clotting and complement activation.
Capsule
Encapsulated strains of bacteria have capsules that consist of high-molecular-weight polysaccharides that form gels and allow bacteria to adhere to the host target cell surface. The capsule is shieldlike because it is hydrophilic and poorly immunogenic. First, the polysaccharide-rich composition of the capsule strongly inhibits phagocytosis by the hydrophobic surface of the host cell. Second, many capsules are poor immunogens45 and complement activators.46 If phagocytosis occurs, it requires specific opsonizing antibodies against the capsule. The thickness of the capsule in Francisella has been correlated with virulence, and in Brucella, encapsulated smooth colonies are ingested less readily than rough colonies.47 The virulence of Klebsiella43 and Yersinia48 species has also been correlated with capsular presence and protection. Immunologic diversity of the capsule within the same species explains the basis for serotyping. Although most ocular isolates of Haemophilus influenzae are nonencapsulated, the encapsulated form of type b is more virulent than the encapsulated type d.49
Flagella and Fimbriae (Pili)
When seen with transmission electron microscopy (TEM), the outer membrane of a gram-negative rod can take on a peritrichous appearance because of the flagella and more numerous pili that surround it. Flagella are larger than pili (they measure 16 to 18 nanometers in diameter) and allow for motility of the organism toward and within the host tissue. Ninety-five percent of clinical isolates of Pseudomonas are flagellated,50 and the burned-mouse model of P. aeruginosa has shown that flagella-deficient mutants are significantly less virulent.51 Flagellar proteins have been studied as vaccines. Rudner and associates52 showed that systemic or topical immunization of mice with strain-specific flagellar proteins or antiflagellar antibody homologous to the specific strain of bacteria protected them from pseudomonal keratitis.
TEM also reveals fimbriae, also known as pili, that measure 4 to 10 nanometers in diameter. These microfibrils can vary in number (2 to 12 in Pseudomonas) and can be classified according to their function as adhesins, lectins, evasins, aggressins, and sex pili. Fimbriated cells adhere to surfaces with specificity and thus allow colonization of a specific host tissue cell. Attachment to host cells is a first step in Pseudomonas keratitis. Stern and colleagues53 demonstrated that murine corneal trauma predisposes to ulceration, not by increasing the exposed area of de-epithelialized stroma for entry of the organisms, but by providing an injured epithelial edge to which P. aeruginosa can adhere. Purified Pseudomonas pili were found to compete with whole bacteria in saturating the binding receptors of host tissue.54 Knutton and coworkers,55 in their experiments with enterotoxigenic E. coli, showed that before invasion, these microorganisms adhere to the mucosal surface of intestinal cells with pili that allow them to hold on to specific mucosal receptors.
Proteolytic Enzymes
Exotoxin A,56 alkaline protease,57 exoenzyme S,58 phospholipase C,59 hemolysin,60 and elastase61 have all been implicated as players in the pathogenesis of Pseudomonas infections. Whereas endotoxins are part of the cellular wall, exotoxins (also known as cytolysins) are extracellular enzymes and are easily separated from the envelope. Most of the cytolysins that have been isolated and purified are made up of two components. One part binds to the target cell and allows the other part, the enzyme, to pass through the cell membrane. After preparing a specific combination of amino acids and a metal-chelating moiety as an inhibitor to Pseudomonas elastase (a zinc metalloendopeptidase), Kessler and associates61 demonstrated that intrastromal injection of the inhibitor first, followed by elastase, prevented corneal melting. Further, subconjunctival injections of the elastase and inhibitor only delayed corneal melting, suggesting that the reversal of corneal melt by the elastase is better carried intrastromally. Purified P. aeruginosa hemolysin injected into the corneas of rabbits was found to induce an extensive leukocytic invasion of the corneal stroma.60 In Moraxella angular conjunctivitis, the pathogenesis for lid maceration is caused by proteases from inflammatory cells rather than proteases elaborated by Moraxella.62 Both encapsulated and untypeable isolates of H. influenzae produce an IgA protease that degrades the protective secretory IgA elaborated by host mucosal surfaces.63,64
BACTERIAL RECEPTORS AND HOST OCULAR SURFACE
Host Surface Glycoproteins and Glycolipids
Adherence of bacteria to the corneal epithelium is a prerequisite for keratitis. Both P. aeruginosa and Staphylococcus aureus were found to bind to rabbit corneal epithelial cells in vitro. P. aeruginosa bound, in multiple layers, to the periphery of cells grown on glass slides, whereas S. aureus bound more randomly to the cell surface. E. coli did not bind significantly to those cells. The peripheral location of Pseudomonas binding is probably caused by its affinity to macromolecules of the cell surface involved in cell-cell interaction.65 Panjwani and coworkers66 also demonstrated that P. aeruginosa binds to rabbit corneal neutral glycosphingolipids. They later demonstrated that P. aeruginosa also binds to specific phospholipids (phosphatidylinositol and phosphatidylserine) extracted from rabbit corneal epithelium.67 These two molecules in ocular mucus or at the corneal surface may function as bacterial receptors and allow specific host-bacterium interaction and initial colonization.
Pseudomonas pili also bind to corneal epithelial receptors, which are glycoproteins.68 More specifically, such carbohydrate receptors have been studied and include sialic acid,69,70 N-acetylmannosamine,71 mannose,72 galactose,73 N-acetylglucosamine,74 and L-fucose.75 Although rabbit and human corneal epithelial cell models were not able to show detectable levels of the glycolipid asialo GM1,76 the incidence of murine P. aeruginosa keratitis was significantly reduced after mouse corneas infected with Pseudomonas were treated with a serum containing antibodies specific to asialo GM1, a glycolipid to which bacterial pili and LPS usually bind. This experiment, performed by Hazlett and associates,77 provides evidence that antibodies against host corneal receptors can significantly inhibit bacterial binding in vitro. In addition, when applied topically in vivo, the antibodies did confer some immunity and decreased the severity of the disease.
Endocytosis
In addition to the traditional extracellular pathogenesis of P. aeruginosa keratitis, the virulence of the infection has more recently been linked to an additional process that is intracellular. In those experiments, P. aeruginosa was seen inside keratocytes of athymic nude mice. TEM revealed that the invaded cells were present in membrane-bound vesicles, suggesting an endocytic process. At 24 hours, many of the bacteria were found free in the cytoplasm.78 In addition to the latter in vivo experiments, P. aeruginosa was found to invade several in vitro models. A temperature of 37°C allowed for a 10-fold increase in invasion of P. aeruginosa compared with experiments performed at 4°C. Further, host cell actin microfilaments may be required in the invasion, because cytochalasin D inhibited invasion.79
BIOFILM FORMATION
In suboptimal environmental conditions, as when nutrients are low, bacteria divide at a slower rate and behave differently. They secrete and live in an extracellular polymer matrix, also known as biofilm.80 Bacteria within a biofilm are relatively protected from the host's immune system and antibiotics and cause chronic and relentless infections. Biofilms are clinically important wherever biomaterials are used because they create a type of surface colonization.
Contact Lenses and Biofilm
In ophthalmic practice, contact lenses and their cases are the most commonly used biopolymers. Since the advent of soft contact lenses, bacterial keratitis has occurred in association with infected cases and contact lens solutions,81,82 associated with poor patient compliance. Pseudomonal keratitis has also been linked to the use of saline soaking solutions prepared from distilled water and sodium chloride tablets.83 Extended overnight contact lens wear is a major risk factor for corneal ulcers,84–86 and among gram-negative rods, Pseudomonas and Serratia species have been the most common culprits (Fig. 1).87,88
|
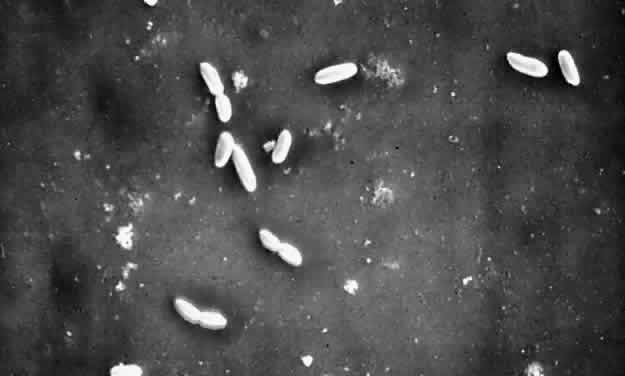
P. aeruginosa is well adapted to grow in aquatic environments such as swimming pools, bathtubs, taps, and bottled and distilled water. Despite disinfectants that kill the single (planktonic) pseudomonal cells, some organisms adhere to the plastic, forming a biofilm. With time, the biofilm releases more planktonic cells, which can divide and replicate in a weaker disinfectant. Serratia species biofilm has been identified on the internal surface of plastic containers,89,90 and the microorganisms have been able to survive in the presence of benzalkonium chloride and 2% chlorhexidine. The ability of Serratia to survive chlorhexidine is attributed to changes in its outer bacterial cell membrane and increased adherence to polyethylene,91 a polymer from which plastic bottles and contact lens cases are manufactured.
Contact Lenses and Mucosal Immunity
The role of mucosal immunity in contact lensassociated bacterial keratitis may shed more light on one aspect of the underlying pathogenesis. Secretory IgA (sIgA) is the most predominant immunoglobulin in tears and plays an important role in preventing bacterial adherence to mucosal tissues.92 Hazlett and colleagues93 showed that sIgA inhibits binding of P. aeruginosa to cornea in a murine keratitis model. Further studies have revealed that the antipseudomonal response of sIgA was significantly lower in extended contact lens wearers than in controls. The mechanism for the lower response remains unclear. One possible explanation is that extended contact lenses cover the cornea and limbal conjunctiva, thus creating a barrier and reducing the chance of antigen presentation to the corneal antigen-presenting cells thought to be important in initiating the IgA response.94
Other Biomaterials
Gram-negative rod infections have been associated with other ophthalmic biopolymers such as ophthalmic sutures,95 intraocular lenses,5 sponges,96,97 and ganciclovir implants98 by means of an adhesive biofilm formation. Although gram-negative rods are isolated less commonly than gram-positive bacteria as causative organisms of suture abscesses, they can lead to endophthalmitis and loss of vision. A biofilm can develop around sutures, especially if loose. Because a suture abscess can lead to endophthalmitis after penetrating keratoplasty, all eroded (broken and loose) sutures should be removed as soon as they are identified.99,100 In vitro studies have demonstrated that P. aeruginosa adheres significantly better to intraocular lenses (IOLs) than does S. aureus (p < 0.05).101 Further, similar studies were performed with heparin-coated IOLs, and the results suggested that there is less P. aeruginosa attachment to the lens, and heparin may reduce the adherence by placing a highly hydrated layer between the bacteria and the IOL surface. The use of heparin-coated IOLs could diminish the incidence of endophthalmitis.102
Because new biomaterials continue to be introduced, research will continue to address ways to prevent biofilm formation and subsequently to reduce sight-threatening bacterial infections.
HYPOXIA
Contact Lenses and Oxygen Transmissibility
Corneal hypoxia has been linked to extended contact lens wear. The oxygen permeability, Dk, measures the amount of oxygen that can diffuse in a material. The oxygen transmissibility (Dk/L) determines how much oxygen a cornea receives after a contact lens of thickness L is placed on it. Overnight wear of contact lenses with poor oxygen transmissibility, especially those with Dk/L total values less than 50 × 10-9 (cm/sec)(ml O2/ml mm Hg), increases the risk of P. aeruginosa keratitis.103 Epithelial damage produced by contact lenses with low oxygen transmissibility worn overnight was shown to be the greatest risk factor for bacterial binding, not lens rigidity. Aswad and colleagues104 demonstrated that the extent of P. aeruginosa binding to extended-wear soft contact lenses was correlated with the number of focal deposits on the lenses, and that P. aeruginosa adhered more avidly to the focal deposits than did S. aureus. TEM further revealed that most of the binding of P. aeruginosa was to large focal deposits (more than 150 μm). These findings suggest that the corneal deposits may serve as points of attachment.
Lid Closure as a Risk Factor for Corneal Ulceration
Both clinical86 and experimental105 studies have suggested that the closed eye constitutes an important risk factor for infection when an extended-wear lens is worn. Aswad and associates105 found a significantly greater incidence of bacterial keratitis in rabbit eyes that had undergone lid closure after the placement of a contact lens as opposed to open eyes with a soft contact lens.
OTHER HOST FACTORS
Dry Eyes and Vitamin A Deficiency
Bacterium-host interaction is a key step in pathogenesis. In a P. aeruginosa keratitis model in New Zealand White rabbits and Wistar rats, Fleiszig and coworkers106 studied the effect of endogenous ocular surface mucus and found that its mucin fraction interfered with the adherence of the bacteria to intact corneal epithelial cells. The authors suggested that abnormal or reduced ocular mucus, as in dry eye conditions, may play a role in promoting P. aeruginosa keratitis.
Trauma and Immune Status
A variety of other host factors, including the immune status, can enhance gram-negative rod virulence. Trauma to the host surface allows easier bacterial adherence and penetration through the injured surface.53 Neutropenic and C3-depleted animals have been shown to have fewer polymorphonuclear leukocytes in the infected cornea and more fulminant infection.107–109
Vitamin A deficiency is associated with dry eyes and xerophthalmia. This represents a leading cause of blindness in developing countries. DeCarlo and associates110 studied pseudomonal keratitis in a vitamin A-deficient rabbit model and found that rabbits with severe xerophthalmia and epithelial keratinization had severe keratitis compared with those with initial mild punctate epithelial keratitis. Lack of mucus, as in certain forms of dry eye, has been experimentally shown to improve bacterial adherence to host tissue.106