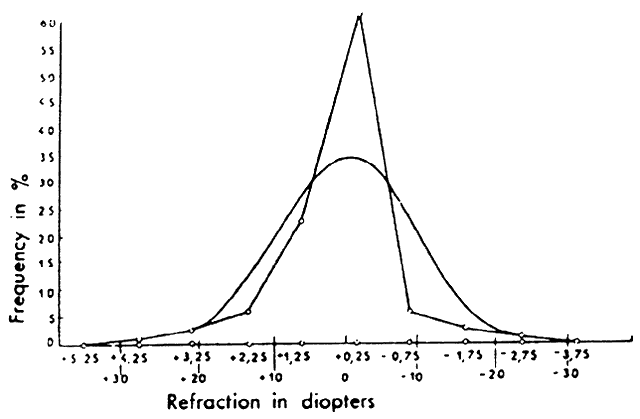
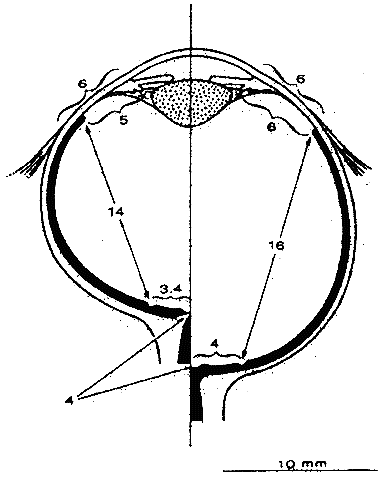
1. Hirschberg J: The History of Ophthalmology, pp 159–160. Blodi F (trans). Bonn, JP
Wayenbough, 1982 2. Kepler J: Dioptrice: Seu demonstratio eorum quae visui et visibilibus propter
conspicilla non ita pridem inventa accidunt. Augsburg, 1611 3. Worth C: Hereditary influence in myopia. Trans Ophthalmol Soc UK, 26: 141, 1906 4. Wold KC: Hereditary myopia. Arch Ophthalmol 42:225, 1949 5. Steiger A: Die Entstehung der Sphärischen Refraktionen des Menschlichen
Auges. Berlin, S Karger, 1913 6. Tron E: The optic elements of the refractive power of the eye. In Ridley
F, Sorsby A (eds): Modern Trends in Ophthalmology. London, Butterworth & Co, 1940 7. Stenstrom S: Variations and correlations of the optical components of the
eye. In Sorsby A (ed): Modern Trends in Ophthalmology. New York, Paul
C. Hoeber, 1948 8. Sorsby A, Benjamin B, Davey JB et al: Emmetropia and its Aberrations, a
Study in the Correlation of the Optical Components of the Eye. Medical
Research Council Report No. 293, London, Her Majesty's Stationery
Office, 1957 9. François J: Heredity in Ophthalmology, pp 186–204. St, Louis, CV
Mosby, 1961 10. Scheerer R: Zur Entwicklung der Brechzustande des Auges. Dtsch Ophthalmol Ges 47:118, 1928 11. Betsch A: Uber die menschliche Refraktionskurve. Klin Monatsbl Augenheilkd 82:365. 1929 12. Franceschetti A: Refraktion. In: Schieck and Brückner's Handbook, vol 1, pp 716–722, 1930 13. Straub M: Uber die Aetilogie der Brechungsanomalien des Auges und den Ursprung der
Emmetropie. Graefes Arch KIln Exp Ophthalmol 70:130, 1909 14. Hartridge H: The causes of errors of refraction. Br J Physiol Optics 7:143, 1950 15. Van Alphen GWHM: On emmetropia and ametropia. Ophthalmologica 142(suppl):l, 1961 16. Michaels DD: Visual Optics and Refraction, a Clinical Approach, pp 457–483. St. Louis, CV
Mosby, 1985 17. Sorsby A, Leary GA: A Longitudinal Study of Refraction and its Components
During Growth. Medical Research Council Report No. 309, London, Her
Majesty's Stationery Office, 1970 18. Sorsby A: The nature of spherical refractive errors. In Sorsby A (ed): Modern
Ophthalmology, vol 3, Philadelphia, JB Lippincott, 1972 19. Rabin J, Van Sluyters RC, Malach R: Emmetropization: A vision-dependent phenomenon. Invest Ophthalmol Vis Sci 20:561, 1981 20. Wixson RJ: Refraction pedigrees: Comments on heredity and environment. Am J Optom 39:471, 1962 21. Arner RS: An evaluation of the Massachusetts vision screening test and its implication
in a genetic theory of myopia. Am J Optom 35:470, 1958 22. Wixson RJ: Statistical analysis of hereditary factors in ametropia. Am J Optom 33:374, 1956 23. Holm S: Les de la refraction oculaire chez les paleonegrieles au Gabon
Afrique Equatoriale François. Acta Ophthalmol 13(suppl): Vol 15, 1937. In
Borish I: Clinical Refraction pp 5–148. Chicago, The
Professional Press, 1975 24. Harris RB: Refractive error in monozygotic twins. Eye Ear Nose Throat Monthly 36:41, 1957 25. Waardenberg PJ: On emmetropia and ametropia. Acta Ophthalmol 142(suppl), 1961. In
Borish I: Clinical Refraction, pp 5–148. Chicago, The
Professional Press, 1975 26. Bays JA: Heredity and myopia. J Am Optom Assoc 37:46, 1966 27. Wixson RJ: The relative effects of heredity and environment upon the refractive
errors of identical twins, fraternal twins, and like-sex siblings, Am
J Optom 35:346, 1958 28. Ditmars DL: A Comparative study of refractive errors of young myopes and
their parents: Am J Optom 44:448, 1967 29. Hirsch M J, Ditmars DL: Refraction of young myopes and their parents: A reanalysis. Am J Optom 46:30, 1969 30. Beyer CK: Heredity and refraction errors, refraction in children. International Ophthalmology Clinics 2:935, 1962 31. Ellerbrock VJ: Developmental, Congenital and Hereditary Anomalies of the
Eye. In Hirsch M J, Wick W (eds): Vision of Children. Philadelphia, Chilton
Book Co, 1963 32. Czellitzer C: Totalrefraktion und Hornhautrefraktion mit besonder Berunksichtigung des
physiologischen linsen Astigmatismos. KIln Monatsbl Augenheilkd 79:301, 1927 33. Kanefuji M: Correlation among refractive errors. Acta Soc Ophthalmol Jpn 57:85, 1953 34. Borish I: Clinical Refraction, pp 5–148. Chicago, The Professional
Press, 1975 35. Jablonski W: Ein Beltrag zur Vererbung der Refraktion menschlicher ugen. Arch Augenheilkd 91:308, 1922 36. Waardenberg PJ: Twin research in ophthalmology. Doc Ophthalmol 84:593, 1950 37. Weekers R, Moaurian P, Harcourt J, Andre A: A contribution to the subject of the genesis of ametropia by the study
of uni- and bi-ovular twins. Arch Ophthalmo1 Fr 17:38, 1957 38. Otsuka Z: Refraction of twins. Acta Soc Ophthalmol Jpn 46: 1744, 1942 39. Orth H: Extreme Diskordanz der Refraktinswerte eineiiger zwillinge. Klin Monatsbl Augenheilkd 124:304, 1954 40. Lisch K: Zur Frase der Myopiegenese. KIln Monatsbl Augenheilkd 106:695, 1941 41. Jancke G, Holste A: Der Einfluss von Erblichkeit und Umwelt auf die Refraktionsentstehung: Eine
Zwillings-studie. KIln Monatsbl Augenheilkd 107:373, 1941 42. Sorsby A, Sheridan M, Leary GA: Refraction and its Components in Twins. Medical
Research Council Report No. 303, London, Her Majesty's Stationery
Office, 1962 43. Heinonen O: Über die Refraktion bei eineiigen Zwillingen, speziell in Hinsicht
der asymmetrischen Fälle. Acta Ophthalmol 2:35, 1924 44. Siemens HW: Die Zwillingspathologie: Zur Aetiologie des Turmschädels nebst mitteilung
einer dermatologischen Methode zur Diagnose der Eineiigkeit bei
Zwillingen. Virchows Arch Pathol Anat 253:746, 1924 45. Beckershaus F: Über eineiige Zwillinge. Z Augenheilkd 59:264, 1926 46. Schmidt WA: Ophthalmologische untersuchungen bei ein- und zweieiigen Zwillingen. Klin Monatsbl Augenheilkd 80:553, 1928 47. Waardenberg PJ: Refraktion und Zwillingforschung. Klin Monatsbl Augenheilkd 84:593, 1930 48. Huber A: Ophthalmologische untersuchungen an ein-eiigen und zweieiigen Zwillingen. Arch Klaus-Stift Vererb Forsch 6:41, 1931 49. Oberhoff K: Hyperopischer Astigmatismus und Hyperopie bei eineiigen Zwillingen. Klin Monatsbl Augenheilkd 107:577, 1941 50. Bücklers M: Hochgradige Myopie bei eineiigen Zwillingen. Klin Monatsbl Augenheilkd 102: 144, 1939 51. Hofmann WP, Carey ET: Congenital myopic astigmatism in identical twins. Am J Ophthalmol 25: 1495, 1942 52. Via|lefont H: Symétrie en miroir, pour un oeil seulement, d'un astigmatisme myopique
chez deux jumelles. Bull Soc Ophthalmol Fr 1:101, 1949 53. Spengler E: Ist Hornbautastigmatismus vererblich? Klin Monatsbl Augenheilkd 42: 164, 1904 54. Steiger A: Studien über die erblichen Verhäaltnisse der Hornhautkrümmung. Z
Augenheilkd 17:307,444, 1907 55. Leibowicz M: Hornhautkrümmung und Astigmatismus. Z Ophthalmol Opt 16:33, 1928 56. Czellitzer A: Der Hornhautastigmatismus, seine Darstel-lung und seine Vererbung. Proceedings
of the International Congress on Ophthalmology, Amsterdam, September 1929. Klin Monatsbl Augenheilkd 83:613, 1929 57. Halbertsma KTA: Untersuchungen bei mehrfachen Zwillingspaaren in derselben
Familie. Proceedings of the 121h International Congress on Ophthalmology, Amsterdam, September 1929, pp 272–277 58. Motolese A, Berardi M: Hornhautastigmatismus und Vererbung. Graefes Arch Klin Exp Ophthalmol 136:27, 1936 59. Gasser I: Über Vererbung extremer Hornhautkrümmung, thesis. Zürich, 1938 60. Bir6 I: Data concerning the heredity of astigmatism. Ophthalmologica 115: 156, 1948 61. Powell S: Data on the heredity of astigmatism. Optician 116:286, 1948 62. Vogt A: Zur Genese der sphärischen Refraktionen. Ber Dtsch Ophthalmol Ges 44:67, 1924 63. Rieger H, Thums K: Die “lehre” Steigers und die Frage der Erbbedingtheit der Refraktionszustände. Graefes Arch Klin Exp Ophthalmol 144:470, 1942 64. Rieger H, Thums K: Augenärztliche untersuchungen im Rahmen des anthropologischerbbiologischen
Vater-schaftnachweises. Wien Med Wochenschr 101:594, 1951 65. Zeki SM: Optic nerve hypoplasia and astigmatism: A new association. Br J Ophthalmol 74:297, 1990 66. Punnett HH, Harley RD: Genetics in pediatric ophthalmology. In Harley RD (ed): Pediatric
Ophthalmology, pp 28–81. Philadelphia, WB Saunders, 1983 67. Kardo Sysoeff W: Zur Lösung des Refraktionsproblems vom Standpunkte
der Biologie. Russ Ophthalmol J 5:689. In: Klin Monatsbl Augenheilkd 77:728, 1924 68. Waardenberg PJ: Total refraction and the variability of its individual
components. In Waardenberg PJ, Franceschetti A (eds): Genetics and Ophthalmology, pp 1201–1285. Oxford, Blackwell Scientific Publications, 1963 69. Scherenberg K: Beiträge zur Lehre vom reinen Mikrophthalmus, thesis. Tübingen, 1900 Nagels Jber, 297, 1900 70. Schneidemann TB: High hypermetropia. Ophthalmol Rec 15:468, 1906 71. Leber T: Über höchstgradige Hypermetropie bei Vorhandensein der Krystallinse. Z Augenheilkd 16:286, 1906 72. Clausen W: Hypermetropia permagna (demonstration). Z Augenheilkd 25:113, 1911 73. Vogelsang K: Excessive Hypermetrie bei Konsanguinität der Eltern. Klln Monatsbl Augenheilkd 99:539, 1937 74. Cavallacci G: Ipermetropia famigliare di alto grado. Arch Ottalmol 44: 172, 1937 75. Scardiaccione M: Sopra una malformazione oculare congenita caratterizzata da pseudopapilla
da stasi con membrana epipapillare, ipermetropia elevata e formazioni
verrucoidi del fondo. Boll Ocul 19:703, 1940 76. Lambert RK, McDonnald CE: Hereditary high hyperopia. Am J Ophthalmol 3: 14, 1931 77. Czellitzer A: Die Vererbung von Augenleiden. Berl Klin Wochenschr 44:2070, 1912 78. Roy FH: Ocular Syndromes and Systemic Disease, p 338. Orlando, FL, Grune & Stratton, 1985 79. Spitznas M, Gerhe E, Bateman JB: Hereditary posterior microphthalmos with papillomacular fold and high hyperopia. Arch Ophthalmol 101:413, 1983 80. Dagi LR, Leys MJ, Hansen RM, Fulton AB: Hyperopia in complicated Leber's congenital amaurosis. Arch Ophthalmol 108:709, 1990 81. Wagner RS, Caputo AR, Nelson LB, Zanoni D: High hyperopia in Leber's congenital amaurosis. Arch Ophthalmol 103:1507, 1985 82. Brodsky MC, Keppen LD, Rice CD, Ranells JD: Ocular and systemic findings in the Aarskog (facial-digital-genital) syndrome. Am J Ophthalmol 109:450, 1990 83. Lin LK, Chen C: Twin study on myopia. Acta Genet Med Gemellol 36:535, 1987 84. Goldschmidt E: On the Etiology of Myopia. Copehagen, Munksgaard, 1968. In
Curtin B J: The Myopias: Basic Science and Clinical Management. Philadelphia, Harper & Row, 1985 85. Karlsson JL: Concordance rates for myopia in twins. Clin Genet 6: 142, 1974 86. Otsuka J: Studies on the influence of heredity and environment on ocular refraction
of the eye. Acta Soc Ophthalmol Jpn 47:889, 1943 87. Otsuka J: The observation of the myopic fundus changes in twins. Acta Soc Ophthalmol Jpn 52:16, 1948 88. Otsuka J: The result of eye and refraction examination of 300 sets of twins. Acta Soc Ophthalmol Jpn 64:46, 1950 89. Bloch I: Discussion on heredity of myopia. Klin Monatsbl Augenheilkd 71:764, 1923 90. Schokking CT: Uitbreiding van het Tweelingonderzoek in Nederland, thesis. Leiden, 1931 91. von Rötth A: Ueber Augenhefund von Zwillingen. Klin Monatsbl Augenheilkd 98:636, 1937 92. Bouterwek H: Erhebungen an eineiigen Zwillingspaaren über Erbanblage und Umwelt
als Charakterbildner. Z Menschl Vererb Konstit Lehre 20:265, 1936 93. Dahl EO: The refraction of children. Am J Ophthalmol 19:422, 1936 94. Holste A: Die Refraktionsverhaltnisse bei ein- und zwei-eiigen Zwillingen. Graefes
Arch KIln Exp Ophthalmol 142:467. 1941 95. Krüger ME: Zwillingbefunde im Gau Mecklenburg, thesis. Borna, Leipzig, 1937 96. Weber E: Die Vererbung der Refraktion an hand von Älteren und neueren Zwillingsuntersuchungen. KIln Monatsbl Augenheilkd 107:574, 1941 97. Jancke G: Zur Entstehung der Kurzsichtigkeit. KIln Monatsbl Augenheilkd 106:264, 1941 98. Halbertsma KTA: Beobachtungen über die Hornhautkrümmung, Astigmatismus und Refraktion
bei mehrfachen Zwillingspaaren. Graefes Arch Klin Exp Ophthalmol 123:632, 1930 99. Gedda L: Studio dei Gamelli. Rome, Orizzonte Medico, 1951 100. Munch O: Uber Anisometropie bei eineiigen Zwillingen. Ber Z Dtsch Ophthalmol Ges 48: 159, 1922 101. Burns RA: Hereditary myopia in identical twins. Br J Ophthalmol 33:491, 1949 102. Masters S: Strabismus in twins. Am J Ophthalmol 35:1825, 1952 103. Meyer-Schwickerath G: Zwillingsstatistische untersuchung uber den Einfluss von Umweltfaktoren
auf den Myopiegrad. Graefes Arch Klin Exp Ophthalmol 149:695, 1949 104. Wibaut F: Discussion after Waardenberg. Klin Monatsbl Augenheilkd 76: 138, 1926 105. Marchesani O: Hochgradige Myopic bei eineiigen Zwillingen. Klin Monatsbl Augenheilkd 94:97, 1935 106. Grassl R: Dementia praecox bei Zwillingen. Arch Rassen Ges Biol 14:177. 1922 107. Nordlöw W: Fall von spontaner Netzhautablösung bei eineiigen Zwillingen. Acta Ophthalmol 16:579, 1938 108. Steiger A: Uber Erbeinheiten am menschlichen Auge. Z Augenheilkd 34:1, 1915 109. Vogel F, Balthasar G: Über ein männliches eineiiges Zwillingspaar mit angeborenem Nystagmus
und Myopie. KIln Monatsbl Augenheilkd 128:456, 1956 110. Weber E: Die Refraktionsverbaltnisse bei Zwillingen. Klin Monatsbl Augenheilkd 106:78, 1941 111. Curtin B J: The Myopias: Basic Science and Clinical Management. Philadelphia, Harper & Row, 1985 112. W61fftin E: Ein myopischer Stammbarm, der über 8 Generationen genau verfolgt werden
konnte. Z Menschl Vererb Konstit Lehre 29:243, 1949 113. Paul L: Untersuchungen über die erbliche Entstehung der Kurzsichtigkeit. Graefes Arch Klin Exp Ophthalmol 139:378, 1938 114. Jaensch PA: Erbliche Entstehung der Kurzsichtigkeit. Med Klin 1:69, 1939 115. Basu SK, Jindal A: Genetic aspects of myopia among the Shia Muslim Dawoodi Bohras of Udaipur
Rajsthan. Hum Hered 33:163, 1983 116. Fukushita K: Clinical studies on heredity in high myopia. Acta Soc Ophthalmol Jpn 86:239, 1982 117. McKusick VA: Mendelian Inheritance in Man: Catalogs of AD, AR and X-Linked
Phenotypes. Baltimore, Johns Hopkins University Press, 1988 118. Karlsson JL: Evidence for recessive inheritance of myopia. Clin Genet 7:197, 1975 119. Bartsocas CS, Kastrantas AD: X-Linked form of myopia. Hum Hered 31:199, 1981 120. Nakajima A: The interrelationship of optical components of the eye and their hereditability, as
studied by phacometry in twins. Jpn J Clin Ophthalmol 14: 1649, 1960 121. Nakajima A: The hereditability estimates of the optical components of the
eye and their mutual relationship by a new method of measurements on
twins. In: Proceedings of the Second International Congress of Human
Genetics, vol 1. Rome, Instituto G. Mendel, 1961 122. Alsbirk PH: Anterior chamber of the eye: A genetic and anthropological study in Greenland
Eskimos. Hum Hered 25:418, 1975 123. Alsbirk PH: Corneal diameter in Greenland Eskimos: Anthropometric and genetic studies
with special reference to primary angle-closure glaucoma. Acta Ophthalmol 53:635, 1975 124. Alsbirk PH: Corneal thickness: II. Environmental and genetic factors. Acta Ophthalmol 56: 105, 1978 125. Albirk PH: Refraction in adult West Greenland Eskimos. Acta Ophthalmol 57:84, 1979 126. Hegmann JP, Mash AJ, Spivey BE: Genetic analysis of human visual parameters
in populations with varying incidences of strabismus. Am J Hum Genet 26:549. 1974 127. Sorsby A, Leary GA, Fraser GR: Family studies on ocular refraction and its components. J Med Genet 3:269, 1966 128. Young FA, Leary GA, Baldwin WR et al: The transmission of refractive errors within Eskimo families. Am J Optom 46:676, 1969 129. Krill AE: Hereditary Retinal and Choroidal Diseases, vol 2, Evaluation. Hagerstown, MD, Harper & Row, 1972 130. Roy FH: Ocular Syndromes and Systemic Disease, p 352. Orlando, FL, Grune & Stratton. 1985 131. Spallone A: Stickler's syndrome: A study of 12 families. Br J Ophthalmol 71:504, 1987 132. Ortiz de Zarate J: Recessive sex-linked inheritance of congenital external ophthalmoplegia
and myopia coincident with other dysplasias. Br J Ophthalmol 50:606, 1966 133. Salleras A, Ortiz de Zarate J: Recessive sex-linked inheritance of external ophthalmoplegia and myopia
coincident with other dysplasias. Br J Ophthalmol 34:662, 1950 134. Gillum WN, Anderson RL: Dominantly inherited blepharoptosis, high myopia and ectopia lentis. Arch Ophthalmol 100:282, 1982 135. Mollica F, LiVolti S, Tomarchio S et al: Autosomal dominant cataract and microcornea associated with myopia in a
Sicilian family. Clin Genet 28:42, 1985 136. Gordon RA, Donzis PB: Myopia associated with retinopathy of prematurity. Ophthalmology 93: 1593, 1986 137. Levensohn G: Zur Anatomie des kurzsichtigen Affenauges. Graefes Arch Klin Exp Ophthalmol 100: 138, 1921 138. Behr C: Uber Kurzsichtigkeit bei Affen. Klin Monatsbl Augenheilkd 62:412, 1919 139. Marchesani O: Untersuchungen über die Myopiegenese. Arch Augenheilkd 104:177, 1931 140. Young FA: The effect of restricted visual space on the primate eye. Am J Ophthalmol 52:799, 1961 141. Wiesel TN, Hubel DH, LeVay S: Changes of ocular dominance columns in monocularly deprived monkeys. Abstract from XL Cold Spring Harbor Symposium, The Synapse 40:581, 1975 142. Wiesel TN, Raviola E: Myopia and eye enlargement after neonatal lid fusion in monkeys. Nature 266:66, 1977 143. Raviola E, Wiesel TN: Effect of dark-rearing on experimental myopia in monkeys. InveSt Ophthalmol Vis Sci 17:485, 1978 144. Wiesel TN, Raviola E: Increase in axial length of the macaque monkey eye after corneal opacification. Invest Ophthalmol Vis Sci 18:1232, 1979 145. Raviola E, Wiesel TN: An animal model of myopia. N Engl J Med 312:1609, 1985 146. Guyton DL, Greene PR, Scholz RT: Dark-rearing interference with emmetropisation in the rhesus monkey. Invest Ophthalmol Vis Sci 29(suppl):33, 1988 147. Von Noorden GK, Crawford MLJ: Lid closure and refractive error in macaque monkeys. Nature 272:53, 1978 148. Smith EL, Harwerth RS, Crawford MLJ, von Noorden GK: Observations on the effects of form deprivation on the refractive status
of the monkey. Invest Ophthalmol Vis Sci 28:1236, 1987 149. Holden AL, Hodos W, Hayes BP, Fitzke FW: Myopia: Induced, normal and clinical. Eye 2(suppl):S242, 1988 150. Lloyd RIL: Birth injuries of the cornea and allied conditions. Am J Ophthalmol 21:359, 1938 151. Angell K, Robb RM, Berson FG: Visual prognosis in patients with ruptures in Descemet's membrane
due to forceps injuries. Arch Ophthalmol 99:2137, 1981 152. Muramatsu T, Sato A, Tokoro T: Influence of corneal opacities on development of myopia. Folia Ophthalmol Jpn 33:1084, 1982 153. Johnson CA, Post RB, Chalupa LM, Lee TJ: Monocular deprivation in humans: A study of identical twins. Invest Ophthalmol Vis Sci 23: 135, 1982 154. Von Noorden GK, Lewis RA: Ocular axial length in unilateral congenital cataracts and blepharoptosis. Invest Ophthalmol Vis Sci 28:750, 1987 155. Robb RM: Refractive errors associated with hemangiomas of the eyelids and orbit
in infancy. Am J Ophthalmol 83:52, 1977 156. Hoyt CS, Stone RD, Fromer C, Billson FA: Monocular axial myopia associated with neonatal eyelid closure in human
infants. Am J Ophthalmol 91:197, 1981 157. Tscherning M: Studier over Myopiens Aetiologi. Copenhagen, 1882. In Curtin
B J: The Myopias: Basic Science and Clinical Management. Philadelphia, Harper & Row, 1985 158. Tscherning M: Studier uber die Aetiology de Myopic. Graefes Arch Klin Exp
Ophthalmol 29:201, 1883. In Curtin BJ: The Myopias: Basic Science and
Clinical Management. Philadelphia, Harper & Row, 1985 159. Cohn H: The Hygiene of the Eye in Schools. Turnbull WP (trans). London, Simpkin, Marshall & Co, 1886 160. Young FA, Beattie R, Newby FJ et al; The Pullman study: A visual survey
of Pullman school children. Am J Optom 31:111; 192, 1954 161. Nadell MC, Weymouth FW, Hirsch MJ: The relationship of frequency of the use of the eyes in close work to the
distribution of the refractive error in a selected sample. Am J Optom 34:523, 1957 162. Angle J, Wissmann DA: Age, reading and myopia. Am J Optom Physiol Opt 55:302, 1978 163. Richler A, Bear JC: Refraction, near work and education: A population study in Newfoundland. Acta Ophthalmol 58:468, 1980 164. Sorsby A: Race, sex and environment in the development of myopia: Preliminary communication. Br J Ophthalmol 12:197, 1928 165. Post RH: Population differences in visual acuity: A review, with speculative notes
on selection relaxation. Eugenics Q 9:189, 1962 |