|
| Chapter 1 Extraocular Muscles JOSEPH L. DEMER Table Of Contents |
| The extraocular muscles (EOMs) play a vital role in the visual system, providing
both static adjustment of binocular alignment necessary to enable
stereopsis, and also the precise dynamic movements necessary to
acquire and maintain visual targets on the fovea despite ubiquitous motion
of the head and body. Ocular saccades, rapid refixational movements
of the eyes, are the fastest movements made by any part of the body, and
maintained foveal fixations are arguably the most precise movements. Every
aspect of the structure of the EOMs and their associated connective
tissues is specialized for the unique functional requirements
of the oculomotor system, and these structural specializations are of
clinical significance for ocular motility. The EOMs differ from all other
striated muscles in many important respects. Description of ocular kinematics requires definition of a coordinate system of rotations. A convenient coordinate system is that of Fick, specifying the order of rotations as follows: (1) horizontal, about a fixed vertical axis; (2) vertical, about a horizontal axis rotated with the eye; and (3) torsional, about the line of sight that has in turn made the preceding two rotations with the eye. An abducting (external) horizontal rotation moves the line of sight temporally, whereas an adducting (internal) horizontal rotation moves the line of sight nasally. An elevating (sursumduction) vertical rotation moves the line of sight toward the vertex of the head, whereas a depressing (deorsumduction) vertical rotation moves the line of sight toward the feet. An intorting rotation about the line of sight rotates the superior pole of the eye nasally, whereas an extorting rotation about the line of sight rotates the superior pole of the eye temporally. Rotational kinematics of any three-dimensional (3D) object such as the eye are not commutative in the mathematical sense.1 The order of rotations of an object is important in determining the resulting orientation, although for most clinical purposes the rotations of normal eyes can be regarded as effectively commutative. In humans there are six oculorotary muscles, functioning as antagonist pairs. The medial rectus (MR) and lateral rectus (LR) muscles accomplish horizontal eye movements, with the MR accomplishing adduction and the LR accomplishing abduction. The superior rectus (SR) and inferior rectus (IR) muscles act as a vertical antagonist pair, with the SR elevating and the IR depressing the globe. However, the vertical rectus EOMs have additional actions that are not strictly antagonistic, as will be detailed below. The superior oblique (SO) and inferior oblique (IO) muscles act as an antagonist pair essential for torsion of the globe around the line of sight. The SO accomplishes intorsion, the IO extorsion. However, the oblique EOMs have additional actions that are also not strictly antagonistic, as will be detailed below. |
| EMBRYOLOGY OF THE EXTRAOCULAR MUSCLES | |
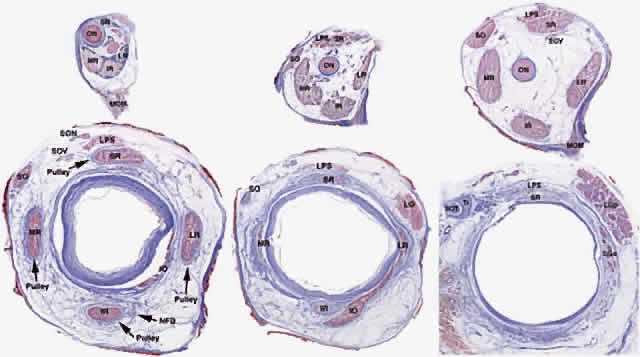
The fibers of the striated EOMs originate from condensations of cranial
mesoderm, whereas their associated connective tissues and the orbital
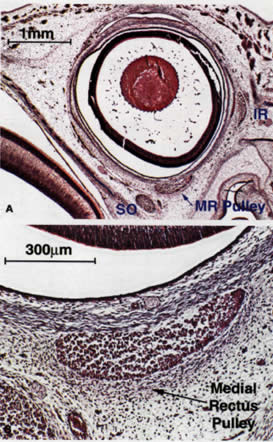
smooth muscles (SMs) originate from the neural crest.2 Myogenesis of the striated EOMs occurs in two waves, primary and secondary.3 Even as early as the primary wave of myogenesis at 11 weeks of gestation (Fig. 1), the perimuscular connective tissues, such as the trochlea and the soft
pulleys of the rectus EOMs, have begun to be associated with the condensations
of myoblasts that will form the EOMs. The periocular connective
tissues develop further by the secondary wave of myogenesis. The
developing EOMs attract innervation from their corresponding cranial
nerves.
|
| INTERNAL STRUCTURE OF THE STRIATED EXTRAOCULAR MUSCLES | |||
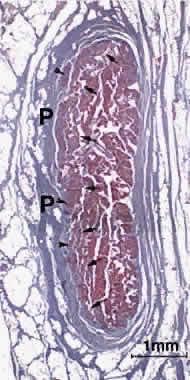
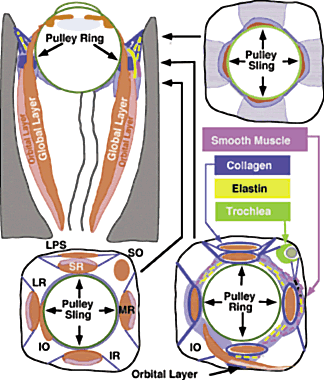
The oculorotary EOMs, although not the levator palpebrae superioris (LPS), consist
of two distinct layers (Fig. 2).4 The global layer, in humans containing a maximum of approximately 10,000 to 15,000 fibers
in the midlength of the EOM, is located adjacent to
the globe in rectus EOMs and in the central core of the oblique EOMs.5 In the rectus EOMs and the SO, the global layer anteriorly becomes the
contiguous tendon and inserts on the sclera.6 In the IO, the global layer inserts directly on the sclera without a tendon. The
orbital layer of each rectus EOM contains 40% to 60% of the
total number of fibers in the EOM, but the number varies according to
the specific EOM; it is greatest for the MR and least for the SR. The
orbital layer of the rectus EOMs and IO does not insert on the globe
at all, but instead inserts on the corresponding connective tissue pulley. The
orbital layer is located on the orbital surface of the rectus
EOM, sometimes forming a C-shaped configuration, and constitutes the
concentric outer layer of the oblique EOMs. An intermediate transitional
zone between the orbital and global layers is sometimes described. For
the global layer of rectus EOMs, the maximum number of fibers is in
midorbit, with the number of fibers diminishing anteriorly and posteriorly.5 This implies that global layer fibers bifurcate and rejoin in myomyous
junctions along the length of each EOM. The number of orbital layer fibers
remains constant along the length of each EOM, suggesting that each
global layer fiber extends the entire length of the EOM.
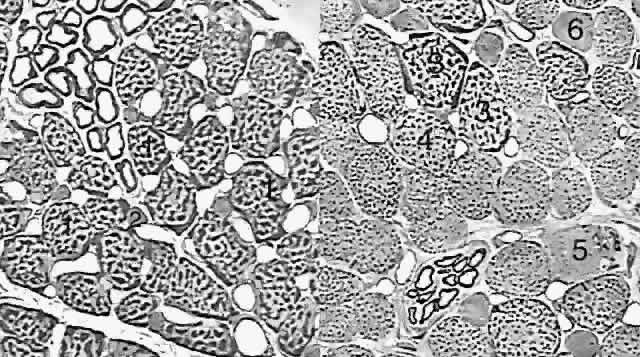
The orbital layer of each EOM contains two distinct muscle fiber types (Fig. 3A).4 About 80% of fibers in the orbital layer of each EOM are fast, twitch-generating, singly innervated fibers (SIFs) resembling mammalian skeletal muscle fibers, whereas 20% are multiply innervated fibers (MIFs) that either do not conduct action potentials or do so only in their central portions.4 Orbital SIFs are relatively small in diameter, and their abundant mitochondria impart a dark staining characteristic with Masson's trichrome. The metabolism and blood supply of orbital SIFs are tailored to their unique mechanical loading and nearly continuous activity. Orbital SIFs are specialized for intense oxidative metabolism and fatigue resistance.4 The vascular supply in the orbital layer is high,2 about 50% greater in humans than the well-perfused global layer. Orbital SIFs express unique myosin isoforms, perhaps related to the requirements of fast twitch capability against continuous loading, because alterations in EOM activity patterns can change EOM-specific myosin heavy chain gene expression.7
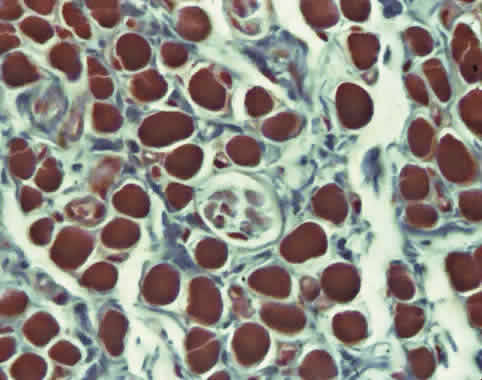
The function of the relatively sparse and primitive orbital MIFs remains unclear. The orbital layer also contains MIFs that either do not conduct action potentials or do so only in their central portions and not near their origins or insertions. The MIFs resemble fibers found in amphibians and are capable of generating slow contractions without twitches. Orbital MIFs may have a proprioceptive function. Also found in the orbital layer are muscle spindles, cylinders of connective tissue containing a handful of small muscle fibers (Fig. 4). Because in skeletal muscles spindles are proprioceptive organs sensing EOM length, the spindles of the orbital layer appear suited to signal the length of the orbital layer and thus the position of the corresponding connective tissue pulley.
The global layer contains one MIF and three designated types of SIFs that really form a continuum distinguished by their density of mitochondria (see Fig. 3B). The largest and most granular SIF is very similar to the orbital SIF, having almost as many mitochondria, whereas the other two SIFs have correspondingly fewer mitochondria. Global MIFs are still smaller, contain fewer mitochondria, and correspond to the orbital MIFs. No spindles are present in the global layer, but the anterior tendonous termination of the global layer of the rectus EOMs contains palisade endings. Palisade endings are distributed along the width of each rectus tendon near the insertion and presumably act as stretch receptors. Like all muscles, EOMs generate tension through the interaction of the proteins actin and myosin. A wide variety of isoforms of myosin are found in EOMs, with the predominant one in orbital SIFs, EOM-specific myosin, being found only in EOMs.4 Myosin isoforms vary along the length of individual EOM fibers. Neonatal and embryonic myosin isoforms persist throughout adult life at the anterior and posterior ends of SIFs. This persistence of immature myosins may be related to the capability of EOMs to adjust their total number of sarcomeres, a process that presumably occurs at the ends of the fibers. Variation in myosin expression may be the reason for the unique susceptibility of the EOMs to certain disorders such as endocrine myopathy, as well as their resistance to other disorders such as muscular dystrophy.8 |
| ORIGINS AND PATHS OF THE STRIATED EXTRAOCULAR MUSCLES | ||
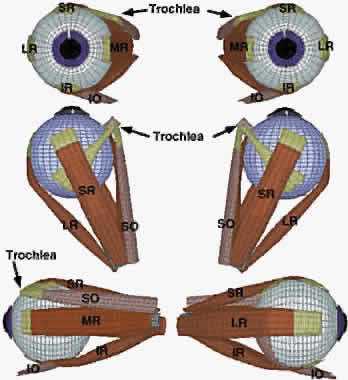
The rectus EOMs originate in the orbital apex from a common fibrous ring
surrounding the optic nerve called the annulus of Zinn (Fig. 5, upper left). The SO muscle originates from the periorbita of the superonasal
orbital wall, slightly more anteriorly (see Fig. 5, upper center). The rectus EOMs course anteriorly through loose, areolar
lobules of orbital fat without mechanical constraint on their paths
until they enter their connective tissue pulleys, described below, that
form sheaths as the EOMs penetrate posterior Tenon's fascia (see Fig. 5, lower left; Fig. 6). The SO muscle remains tethered to the periorbita via connective tissues
as it courses anteriorly, thins to become continuous with its long, thin
tendon (see Fig. 5, lower center), and enters a dense connective tissue sheath as it penetrates
posterior Tenon's fascia posterior to the trochlea (see Fig. 5, lower right). After reflection in the trochlea, the SO tendon passes
beneath the SR, thins, and flattens as it spreads out to its broad scleral
insertion. The IO muscle originates much more anteriorly from the
periorbita of the inferonasal orbital rim adjacent to the anterior lacrimal
crest and runs laterally to enter its connective tissue pulley
immediately inferior to the IR at the point the IO penetrates Tenon's
fascia (see Fig. 5, lower center).
The long axes of the bony orbits are angled about 23 degrees laterally from the midsagittal plane (see Fig. 6). The general configuration of the rectus EOMs and the SO is that of a cone, although it should be clear from Figure 5 that in the posterior orbit this cone is incomplete, without interconnections among adjacent EOMs. As they continue anteriorly, the rectus EOMs thin to become straplike bands about 10 mm in width, and ultimately their global layers become continuous with tendons that insert on the globe. |
| PULLEYS OF EXTRAOCULAR MUSCLES | ||||||||||||
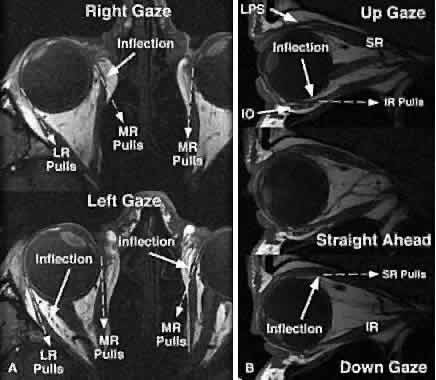
An important insight from contemporary tomographic imaging of EOMs is that
they do not follow straight-line paths from their origins to their
scleral insertions, as implicitly and explicitly (but nevertheless incorrectly) indicated
by older and even many modern textbooks. Discrete
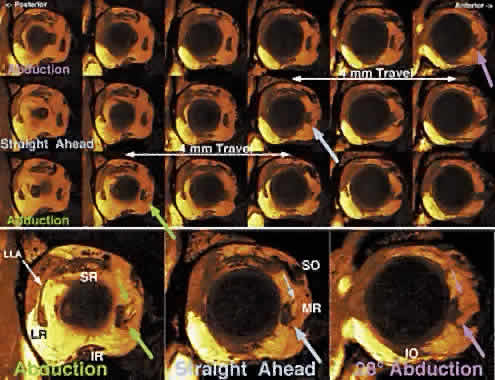
inflections in horizontal rectus EOM paths are obvious on axial images
in abduction and adduction (Fig. 7A). These inflections occur more anteriorly in relaxation of the involved
EOM and more posteriorly during its contraction. The paths of the vertical
rectus EOMs are similarly inflected and are obviously not straight
lines (see Fig. 7B). The rectus EOMs in fact do follow linear paths from the annulus of Zinn
to discrete points in the anterior orbit, paths that are remarkably
stable even during the largest possible shifts of gaze in every direction. The
stability of anterior rectus EOM paths is illustrated in Figure 8 (bottom), where despite substantial ocular abduction, adduction, elevation, and
depression, only the junction of the optic nerve with the globe
is seen to shift relative to the orbit. Posteriorly in the orbit (see Fig. 8, top), the EOMs also maintain stable paths, but the cross sections of
contracting EOMs increase, whereas the those of relaxing EOMs decrease. These
changes in cross section are dynamic indications of rectus contractility, as
well as contractility of the SO and IO.9,10 Atrophic EOMs exhibit a marked reduction in cross section and fail to
demonstrate a contractile increase in cross section in their field of
action.11
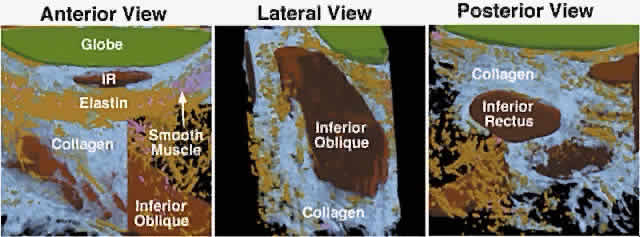
The discrete points in the anterior orbit where the EOM paths are inflected constitute the functional, mechanical pulleys, whose structure will be described in detail below. Anterior to the pulleys, rectus EOM paths shift, sharply when necessary, to follow the scleral insertions in eccentric positions of gaze. The pulleys thus act as functional mechanical insertions of the rectus EOMs. The pulling direction of each EOM is thus defined by the line segment connecting the EOM's scleral insertion to the respective pulley, so that the location of the pulley influences EOM action profoundly. Pulleys consist of discrete rings of dense collagen encircling the EOM and are about 2 mm in length, coaxial with less substantial collagenous sleeves around the EOM (Fig. 9; see Fig. 2). Anteriorly, these sleeves thin to form slings convex to the orbital wall, and more posteriorly the sleeves thin to form slings convex toward the center of the orbit. The anterior pulley slings are also known as the intermuscular septa. The collagenous pulley ring forms the fulcrum of the pulley that inflects the EOM path.
Not surprisingly, given this pivotal mechanical role, electron microscopy demonstrates the fibrils of pulley collagen to have an unusual criss-crossed configuration suited to high internal rigidity.12 Elastic fibers in and around pulleys provide reversible extensibility essential to these resilient tissues.13–15 Reversible extensibility, particularly in connective tissue bands that connect the pulleys to bony anchors on the orbital rim, means that the pulleys are suspended under elastic tension that tends to draw them anteriorly. Bands of SM in the pulley suspensions (see Fig. 9), and particularly the peribulbar muscle connecting the nasal aspects of the vertical rectus pulleys, provide a means of active generation of modulatory force on the pulleys.14 The overall structural organization of the pulleys is illustrated in Figure 10. The anterior pulley slings, forming what has also been termed the intermuscular membrane, are routinely dissected at strabismus surgery. Note that the orbital layers of each rectus EOM insert on their respective pulleys and act to oppose the passive tension produced by the elastic suspensions of the pulleys from the anterior orbit. The pulley suspensions have also been described as check ligaments, a term that fails to describe their mechanical functions clearly. The region of the pulleys and their sleeves is endowed with dense connective tissue that constrains EOM paths. Posterior Tenon's fascia forms a socket for the globe, suspended from the orbital rim like a drumhead. The peripheral part of posterior Tenon's fascia is very dense with collagen, elastin, and SM in circumferential distributions. The pulley sleeves penetrate this part of posterior Tenon's fascia. The central part of posterior Tenon's fascia is thin and compliant, so that the optic nerve, the ciliary nerves, and the ciliary vessels can pass through it easily despite large gaze shifts. Note that posterior to the pulleys, there is no connective tissue to constrain EOM paths. There is no “muscle cone,” only loose, areolar lobules of fat among the EOMs, vessels, and nerves of the posterior orbit. Surgical dissection of the pulley rings or of posterior Tenon's fascia should be avoided, for in addition to liberating deep orbital fat, this will damage the essential function of the pulleys and promote adherence to the globe and scarring of these dense connective tissues that can severely limit normal eye movements.
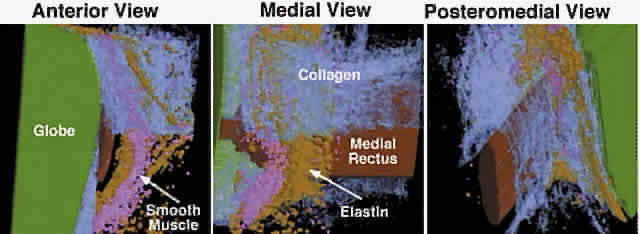
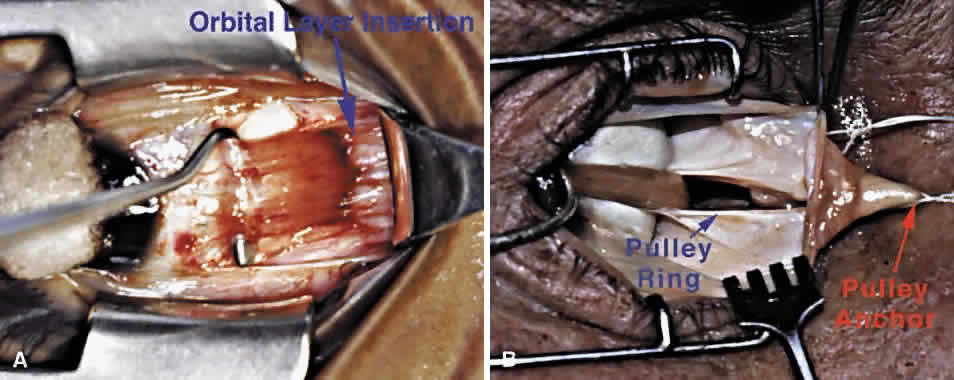
Detailed views of the 3D structure of the rectus pulleys can be obtained by computer-assisted reconstruction of serial, coronal histologic sections stained for specific tissue constituents such as collagen, elastin, and an alpha-actin specific to SM. Such reconstructions, when manipulated using computerized virtual reality techniques, enable orbital tissues to be visualized from perspectives not otherwise possible. Figure 9 shows a posteromedial view of the MR pulley, as if seen from the surgically inaccessible center of the orbit, demonstrating its sleevelike character. Figure 9 also provides a surgically possible anterior view of the MR pulley region, in which the collagen in the inferior half of the pulley has been rendered transparent to display the embedded elastin and the extensive peribulbar SM. The anatomic relations above can, with the benefit of the preceding insights, be readily demonstrated in gross dissections and in surgical exposures. After a typical conjunctival incision and engagement of a rectus muscle on a surgical hook, the white anterior pulley slings come immediately into view. These tissues play little role in constraining EOM paths, although they are related to the more posterior pulley rings. The anterior pulley slings can, with the occasional exception of the lateral levator aponeurosis at the superior border of the LR, be posteriorly displaced by blunt dissection, although sharp dissection is sometimes unavoidable despite its undesirability. After this posterior displacement, fine fibrous bands constituting the insertion of the orbital layer of the EOM on the glistening white pulley suspension can be visualized (Fig. 11A). This insertion site is intimately related to the mechanically effective site of the pulley, the connective tissue ring, as better demonstrated in cadaveric preparations such as Figure 11B. In such preparations, the anterior anchoring point of the MR pulley may be confirmed to be at the anterior lacrimal crest. With detachment of the orbital layer from the MR pulley, the pulley ring may be stretched under high tension to demonstrate the complete ring in posterior Tenon's fascia (see Fig. 11B). Unless the insertion of the orbital layer on the pulley is severed (generally an unnecessary surgical maneuver), the MR cannot through surgical accident slip posteriorly through the pulley to become “lost” in the deep orbit. All the other rectus EOMs are similarly protected against loss by their orbital layer attachments to their pulleys and, in the case of the IR and LR, to the adjacent IO.
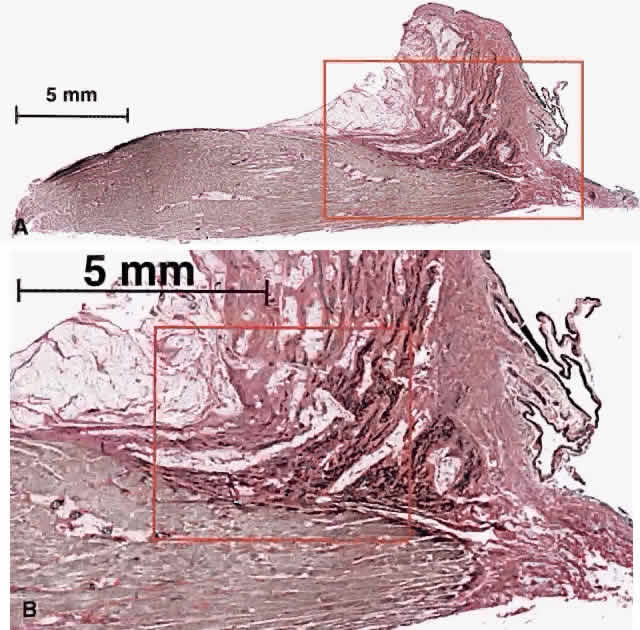
Longitudinal sectioning provides histologic demonstration of the relations of the MR pulley to surrounding tissues (Fig. 12). Even in unstained material, the attachment of the elastin-rich suspension of the MR pulley to the orbital layer of the MR can be seen grossly. Exceptionally dense bands of elastin travel anteriorly from the orbital surface of the pulley to the anterior lacrimal crest and, for other rectus EOMs, the anterior orbital rim. These elastin bands on gross dissection are typically visible as pink striations embedded in the surrounding whiter collagen. Histochemical staining specific for elastin demonstrates dense deposits of elastin running anteriorly from the pulley toward the anterior lacrimal crest. Higher-power microscopy also identifies bands of SM in the pulley suspension as well. These tissues are ideally configured to provide passive elastic force on the MR pulley directed predominantly anteriorly but also to a lesser extent medially.
The dense bands of elastin in the MR pulley suspension insert focally on the orbital surface of the pulley. Because these elastin bands have high contrast with the surrounding collagen on T1-weighted magnetic resonance imaging (MRI), the bands provide a signature to mark the location of the MR pulley in coronal imaging. This MRI signature consists of one to most typically three dark bands iso-dense with muscle extending radially toward the medial orbital wall (Fig. 13). The anteroposterior extent of the orbital layer insertion is sufficiently focal that it is imaged in only one or two adjacent 2-mm-thick coronal planes. Large horizontal gaze shifts cause a corresponding shift in the anteropos-terior location of the MR orbital layer insertion. In 28 degrees of adduction, the MR orbital layer insertion is relatively posterior at the level of the globe-optic nerve junction. In primary gaze, the MR orbital layer insertion moves two image planes (4 mm), and another two image planes (4 mm) an-teriorly in 28 degrees of abduction for a total travel of about 8 mm.
The IR pulley is unique in that it is intimately coupled with that of the IO in a bond that has been termed Lockwood's ligament. In fact, the pulleys of the IR and IO are formed of a common sheath of collagen stiffened by a heavy elastin deposit at the point of crossing.10,16 The orbital layer of the IR inserts on its pulley and does not continue anteriorly. SM and elastin are present in the posterior Tenon's fascia supporting the combined IR/IO pulley, as demonstrated in a computerized reconstruction of this region from serial histologic sections (Fig. 14). The peribulbar SM has its inferior insertion on the nasal aspect of this conjoint pulley and is positioned on contraction to displace the pulley nasally. Inferior oblique contraction in elevation would be expected on anatomic grounds to displace the conjoint IR/IO pulley nasally relative to its position in depression. The SM retractors of the lower eyelid ( Müller's inferior tarsal muscle) and connective tissues extending to the inferior tarsus are also anatomically coupled to the conjoint IR/IO pulley, an arrangement tending to couple lower eyelid position with vertical eye position during depression.
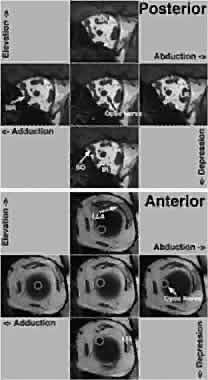
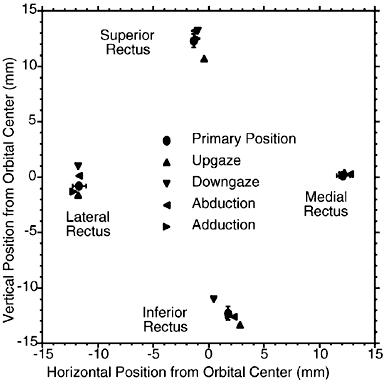
The SR pulley is intimately related to the pulley of the LPS muscle immediately superiorly. The LPS pulley is also a discrete collagenous ring suspended from the superior orbit by Whitnall's ligament and stiffened by a modest amount of elastin and SM, and it is also closely coupled to the adjacent pulley of the SR. In distinction to the oculorotary muscles, the LPS lacks an orbital layer and has only a global layer that passes through its pulley to insert on the anterior border of the densely collagenous tarsal plate. However, posterior motion of the LPS pulley during elevation is achieved by that pulley's intimate mechanical coupling to the SR pulley, which is actively translated posteriorly by its insertion from the contracting SR orbital layer. This arrangement tends mechanically to coordinate upper eyelid position with vertical eye position. The SR pulley also has discrete mechanical couplings to other pulleys. The most prominent such coupling is the lateral levator aponeurosis (LLA), a dense band extending from the lateral border of the conjoint SR/LPS pulley to the superior border of the LR pulley. The LLA contains dense collagen and elastin throughout. The superior third of the LLA contains a striated muscle; the lateral two thirds is richly endowed with SM. The orbital lobe of the lacrimal gland lies on the external surface of the LLA. Although the rectus and IO pulleys are actively mobile along the axes of their respective muscles, rectus pulleys are located quite stably and stereotypically in the coronal plane. Rectus pulley locations may be inferred from the paths of the EOMs in the anterior orbit immediately posterior to the pulley rings. Figure 15 indicates the 95% confidence intervals for the horizontal and vertical coordinates of normal rectus pulleys, showing that they vary by less than ±1 mm.17 This precise placement of rectus pulleys is of obvious importance because the pulleys serve as the functional mechanical origins of the EOMs. Aging, however, may cause small, systematic changes in horizontal rectus positions, which appear to shift downward by 1 to 2 mm from young adulthood to the seventh decade of life (Clark, Miller, and Demer, unpublished data, 1999).
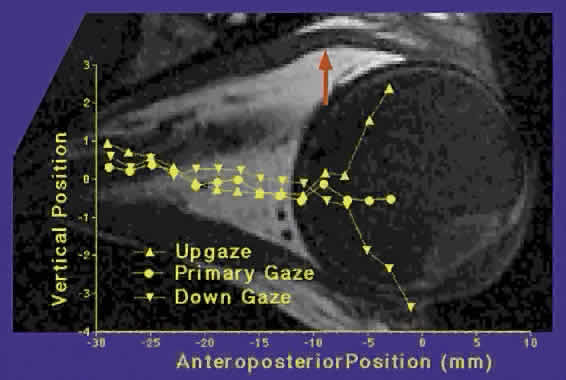
In addition to the precision of the static placement of the pulleys, they are also highly stable during even large gaze shifts (see Fig. 15).17 The MR pulley moves little with horizontal or vertical gaze. The IR pulley, because of its coupling with the IO pulley, is drawn nasally by IO contraction in elevation and moves temporally during IO relaxation during depression by connective tissue elasticity. The SR pulley is relatively stable in the mediolateral direction, for which is well supported by connective tissue bands and Whitnall's ligament running nasotemporally, but moves inferiorly in elevation as it is posteriorly displaced by the SR orbital layer, and superiorly in depression as the SR orbital layer relaxes. The vertical motion of the SR/LPS pulley is transmitted to the LR pulley via the LLA, causing the LR pulley to move superiorly during depression and inferiorly during elevation. The small, gaze-related shifts in rectus pulley positions are also highly stereotyped among normal persons. Recent improvements in MRI now make it possible to image rectus EOM paths anterior to the pulleys, even approaching the scleral insertions, in primary and secondary gaze positions. Reliable measures of rectus EOM paths can be obtained if data are averaged across multiple subjects. Such methodology demonstrates that rectus pulley inflections are discrete and shift with gaze, as suggested by the less rigorous EOM path data above. Figure 16 superimposes on a high-resolution axial MRI through the planes of the horizontal rectus muscles a graph of the vertical position of the LR path as measured at locations throughout the anteroposterior extent of the orbit. The graph, which is horizontally to scale with the MRI image, shows a straight-line path of the LR in primary gaze. However, in elevation and depression, there is a discrete inflection in the LR path at the point corresponding to the LR path inflection seen in the axial MRI image. The LR inflection point moves slightly superiorly in elevation and slightly inferiorly in depression.
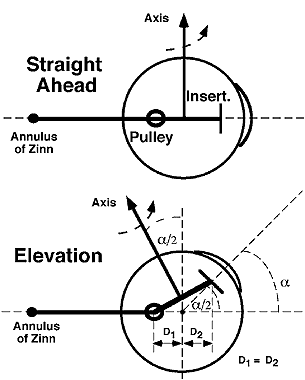
The stereotypic structure and intricate properties of the pulleys suggest that they are important to ocular motility. The pulleys function as the mechanical origins of the EOMs and play a vital role in ocular kinematics, the rotational properties of the eye. Because rotations are not mathematically commutative, final eye orientation depends on the order of rotations.18 Each combination of horizontal and vertical eye positions could, for an arbitrary 3D object, be associated with infinitely many torsional positions.19 The eye is constrained in its torsional freedom (with the head upright and immobile) by a relation known as Donder's law, stating that there is only one torsional eye position for each com-bination of horizontal and vertical eye positions.1 Listing's law, a specific case of the more general Don-der's law, states that any physiologic eye orientation can be reached from any other by rotation about a single axis, and that all such possible axes lie in a single velocity plane, Listing's plane. Listing's law is satisfied if the axis of ocular rotation shifts by exactly one half of the shift in ocular orientation for any eye movement.20 This is the so-called Listing's half-angle rule. Normal pulleys form a natural mechanical substrate for Listing's law and other aspects of ocular kinematics, and they render ocular rotations effectively commutative.21 Figure 17 is a lateral view of a diagrammatic globe showing a horizontal rectus EOM. The rotational axis of the EOM is perpendicular to the line connecting its pulley with the scleral insertion. Thus, the rotational axis of the horizontal rectus EOM is vertical in straight-ahead gaze. Now consider the situation during visual fixation of a horizontally centered target at angular elevation α. If the distance from the pulley to globe center D1 is equal to the distance from the insertion to globe center D2, then the rotational axis will tilt posteriorly by approximately angle α/2, as required by Listing's law.
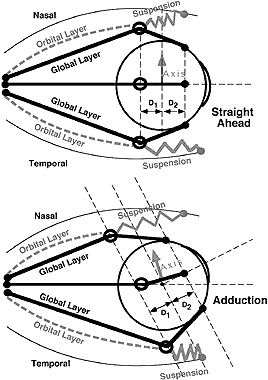
In tertiary (oblique) gaze positions, pulley behavior can still explain Listing's law, as illustrated in Figure 18, a top view of a schematic orbit showing the MR and LR muscles. Starting in primary position in the upper panel, the secondary positions of elevation and depression can be attained in conformity to Listing's half-angle rule, as explained above, if the distance from the pulley to globe center D1 is equal to the distance from the insertion to globe center D2. Starting in adduction, the tertiary positions of adducted elevation and adducted depression can be attained in conformity to Listing's law once again if the distance from pulley to globe center D1 is equal to the distance from insertion to globe center D2. Distances D1 and D2 are referenced to the globe, not relative to the orbit.
To implement Listing's law, shifts in the anteroposterior position of the pulleys relative to the orbit must occur when beginning in the secondary positions of abduction, elevation, or depression, and moving into tertiary positions. Such anteroposterior shifts in pulley positions in secondary gaze positions are readily demonstrated by MRI (see Fig. 13). The SO muscle courses anteriorly from its origin about 40 mm and passes through a U-shaped plate of fibrocartilage, the trochlea, measuring 4 mm wide by 6 mm long.22 With the globe in primary position, the SO becomes tendonous about 10 mm posterior to the trochlea. The trochlea is attached to the superonasal orbital wall about 5 mm posterior to the superior medial angle of the orbital margin and about 10 mm from the medial border of the SR in primary position. A connective tissue sleeve enveloping the SO tendon thickens just posterior to the trochlea and inserts on it, whereas the tendon passes through the trochlea without a true synovial lining. Within the trochlea, the SO tendon has a diameter of only 1 to 2 mm. In primary position, the path of the SO is deflected by the trochlea 54 degrees posterolaterally and passes inferior to the SR just posterior to the scleral insertion of that EOM. The SO tendon expands at roughly the medial border of the SR, flattening and widening to become thin and nearly transparent before inserting on the sclera. |
| SCLERAL INSERTIONS OF THE STRIATED EXTRAOCULAR MUSCLES | |
Anteriorly, the global layers of the rectus EOMs become contiguous with
their tendons, which in turn insert on the sclera of the globe. The insertions
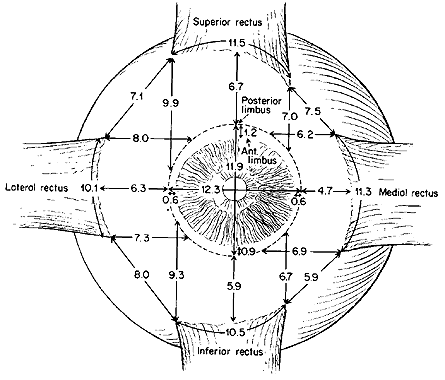
of the rectus EOMs have been described in detail by Apt.23 The width of the rectus EOM insertions varies from the average of 10.1 mm
for the LR to 11.5 mm for the SR; the IR width is 10.5 mm and the
MR width 11.3 mm. The distance between the rectus EOM insertions varies
from the average of 5.9 mm between the MR and IR to 8 mm between the
IR and LR. The distance between the LR and SR muscles is 7.1 mm and between
the SR and MR muscles is 7.5 mm. The location of the scleral insertion
to the corneal limbus varies systematically among the rectus EOMs (Fig. 19); it is shortest for the MR and progressively greater for the IR, LR, and
SR muscles. The progressively increasing distance from limbus to insertion
is known as the spiral of Tilleaux. The lengths of terminal tendons
also vary, from 4 mm for the MR, 6 mm for the IR and SR, and 9 mm
for the LR. Approximately half of the blood supply to the anterior
segment of the eye is transmitted by the rectus EOMs via the anterior
ciliary veins and arteries. Excessive surgical interruption of the anterior
segment blood supply by surgery on multiple adjacent rectus EOMs
can lead to a syndrome of anterior segment ischemia.24
The IO muscle runs laterally from its origin on the orbital plate of the superior maxilla a few millimeters posterior to the inferior orbital rim and adjacent to the lacrimal fossa. Although it is oval at its origin, the IO flattens and follows a straight path temporally to its pulley conjoined with, and inferior to, that of the IR. Lateral to the pulley, the path of the IO curves posteriorly and superiorly at an angle in cadavers of about 75 degrees from the axis of the orbit; this angle varies from 35 to 50 degrees from the visual axis.22 The IO passes medial to the LR, flattening lateral to the pulley to insert without a consistent tendon roughly beneath the fovea along a line angled roughly 25 degrees to the line of insertion of the SO. This insertional line is variable and often includes gross irregularities such as serrations or dehiscences. In 100 specimens, the length of the IO insertion averaged 9.6 mm but varied from 5 to 14 mm.22 The nasal border of the IO insertion averages 4.2 mm lateral to the optic nerve, whereas the temporal border averages 9.5 mm medial to the inferior border of the LR insertion. There is considerable variability in this insertion. The anterior 32 mm of the SO is tendonous. The scleral insertion of the SO is the most variable of all the EOMs.22 The average length of the insertion is 11 mm, but the range in 100 specimens was 7 to 18 mm. The angle of the SO insertion is also markedly variable. On average, the anterior pole of the SO insertion is 4.5 mm posterior to the temporal end of the SR insertion; the SO insertion is roughly linear to a posterior point averaging 6.5 mm from the optic nerve. However, the variability of the SO insertion is so great that it cannot be anticipated with any degree of accuracy by the strabismus surgeon.22 |
| SMOOTH MUSCLES OF THE ORBIT |
| Müller25,26 described SMs in four areas of the human orbit: the inferior palpebral muscle, the superior palpebral muscle, the “orbital muscle” spanning the inferior orbital fissure, and the “peribulbar muscle” surrounding the anterior aspect of the globe. More recently, immunohistochemical staining for human SM alpha-actin has shown that SM cells are especially abundant in the suspensory bands of the pulleys13,14 and may form part of what was described by Müller in the last century as the peribulbar muscle. Despite this similarity, the pulley SM cells are anatomically and structurally distinct from the more anterior palpebral SMs27 and the more posterior orbital muscle of Müller. In particular, the SMs around the MR pulley are organized into bundles, not typical of Müller's palpebral muscle.27 The dense peribulbar SM (see Fig. 10) may play a role in the mechanical configuration of the pulleys to control vergence.28 |
| INNERVATION OF EXTRAOCULAR MUSCLES |
| The majority of the EOMs are innervated by the oculomotor nerve. The superior
division of the oculomotor nerve innervates the SR and LPS, whereas
the inferior division innervates the MR, IR, and IO. The trochlear
nerve innervates the SO, whereas the abducens nerve innervates the LR. The
motor nerve enters each rectus EOM on the orbital surface at about
the junction of the posterior third with the anterior two thirds of
its length. The nerve and blood supply to the IO enters the posterior
portion of the EOM just lateral to the IR, and thus at the lateral aspect
of the combined IO/IR pulley. The neurofibrovascular bundle entering
the IO is compliant over the normal range of motion of the globe, but
after surgical dissection it can be felt to be markedly stiff and
may contribute to the determination of IO path after anterior transposition.29 The trochlear nerve crosses over the SO muscle from the medial side to
enter its upper surface 12 mm anterior to the EOM origin.22 In each case, the portions of EOMs near the myoneural junction contain
luxurious arborizations of myelinated nerves, making up as much as half
of the cross-sectional area of the EOMs in that region. Groups of EOM fibers innervated by a single motor neuron are called motor units. Motor units in the EOMs have the smallest number of fibers of any muscles in the body, reflecting the need for precise control.4 It is estimated that as few as two or three motor units could deviate the eye by 1 degree, and there are some 1100 motor neurons in the cat abducens nucleus.30 There are far more motor units than would be required if all had the same behavior. Some motor neurons in the abducens nucleus project exclusively to either the orbital or the global fiber layer of the LR muscle.31 In cat, the most powerful and fatigue-resistant motor units of the LR, representing 27% of all units, consist of single neurons innervating fibers in both the orbital and global layers.31 These “bilayer” motor units would command similar contraction in the two layers, an arrangement convenient to maintain the position of the pulley relative to the EOM insertion in secondary gaze positions, as required for Listing's half-angle rule. In addition to the fundamental differences in the functions of the orbital and global EOM layers, there is evidence that different motor units may participate differently in different types of eye movements. Details of this complex behavior are currently unclear. The orbital SMs involving the pulley system are autonomically innervated via a complex system of fine, nonmyelinated fibers.14 A sympathetic projection from the superior cervical ganglion travels along the internal carotid artery and orbital vessels and employs norepinephrine, which presumably evokes SM contraction. This action is opposed by the relaxing action of cholinergic, parasympathetic innervation, probably from the ciliary ganglion. Orbital SMs are also relaxed by nitroxidergic parasympathetic innervation employing nitric oxide, a gaseous neurotransmitter found in most SM systems throughout the body. Nitroxidergic projections to the orbital SMs come mainly from the pterygopal-atine ganglion and its extension in the orbital apex, although a few may arise from a small population of nitroxidergic neurons in the ciliary ganglion. |
| PHYSIOLOGY OF EXTRAOCULAR MUSCLES | ||
| Multiply innervated, striated EOM fibers in the orbital layer have twitch
capability near their central end plate bands but generate only graded
contractions proximally and distally along their lengths.4 This behavior is unique among all muscle fiber types. Global MIFs exhibit
slow, graded, nonpropagated responses after either neural or pharmacologic
stimulation. Neither type of MIF is found in mammalian skeletal
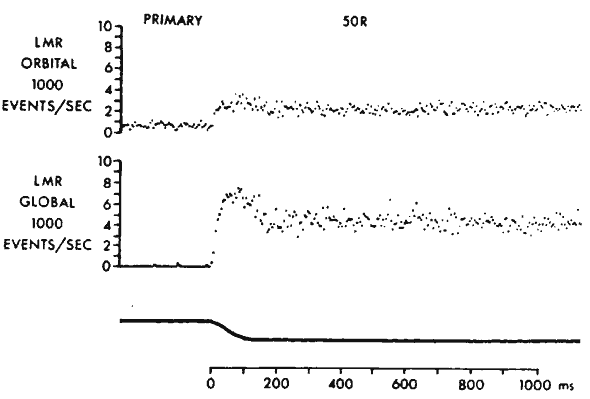
muscle. In contrast to the sluggish behavior of MIFs, SIFs are probably the fastest and most metabolically active muscle fibers in the body.4 The critical fusion frequencies of SIFs are high, with many exceeding 150 Hz.32 Isometric twitch contractions of EOMs are about twice as fast as in skeletal muscles, consistent with the requirements of ocular saccades, the fastest movements made by any part of the body. Orbital singly innervated, and red global singly innervated, fibers have the highest fatigue resistance and oxidative capacity of all EOM fiber types. Blood flow in EOMs is higher than in any skeletal muscles. These observations emphasize that EOMs are fundamentally different from skeletal muscles.4 Differences in EOM fiber types are consistent with different functional roles. As illustrated by the electromyographic (EMG) data in Figure 20, orbital layer fibers have lower recruitment thresholds than global layer fibers.33–35 Many global layer fibers are quiescent just out of their fields of action, whereas orbital layer fibers are generally active throughout the oculomotor range and have higher EMG activity throughout. This behavior is required by the mechanical properties of the orbital tissues. The mechanical load on the orbital layer consists mainly of the elasticity of the attached pulley suspension. Collins has pointed out that the main load on an EOM attached to the globe is viscosity arising from the relaxing antagonist EOM.34 Thus, during saccades, orbital layer fibers develop an initial phasic pulse of activity to move the globe briskly despite the viscosity of the relaxing antagonist, followed by a tonic step of activity to maintain the new globe position against the lesser static elastic load (Fig. 21). A phasic pulse of force in the orbital layer is unnecessary to achieve brisk pulley motion against the mainly elastic load of the connective tissue suspensions of the pulleys, accounting for the finding of only step changes of EMG activity during saccades in the orbital layer. However, elastic loading by passive connective tissue requires that orbital layer fibers maintain active tension throughout the oculomotor range to avoid slack (zero tension) out of the field of EOM action. In contrast, global layer fibers remain under tension even when not actively innervated, because the antagonist EOM stretches them. This latter property allows global layer fibers to exhibit initial electromyographic activity farther into their field of action than global layer fibers, and the greater resting activity in the orbital layer accounts for its greater blood flow and oxidative capabilities.
|
| COOPERATIVE ACTION OF EXTRAOCULAR MUSCLES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tension in each EOM arises from two sources: (1) passive elastic force
arising from the stretching of the EOM by external forces such as those
generated by other EOMs and connective tissue elasticity; and (2) active
contractile force resulting from the energy-dependent interaction
of actin and myosin.36 Passive elastic force, arising from EOM stretching, is directly related
to EOM length and is strongly influenced by strabismus surgery. The
relation between EOM tension and EOM length is a two-dimensional surface
also influenced by innervation of the EOM.36 One strengthening action of surgical resection, usually just of the tendon, of
an EOM is due to stretching the remaining EOM so that its passive
elastic force is increased. Conversely, one weakening action of recessing
the insertion on the sclera of an EOM is due to slackening of
the EOM to reduce its passive elastic force. Active contractile force
is commanded by innervation but also depends on EOM length. The relation
of active EOM tension to innervation and length is also a 3D surface.37 Although strabismus surgery cannot directly change innervation, it can
change the location on the length-tension-innervation surface at which
the innervation is applied, and thus can influence the tension that
innervation will develop. Thus, another effect of resection is to move
the EOM's operating point higher on the length-tension-innervation
surface, increasing EOM tension. Another effect of recession is to
move the EOM's operating point lower on the length-tension-innervation
surface, reducing EOM tension. The effects of neither recession
nor resection directly change innervation or immediately alter the number
of sarcomeres. Antagonist EOM pairs interact mechanically, mutually
establishing the operating points on their length-tension-innervation
surfaces. It should be apparent from the preceding analysis that even the actions of the simplest antagonist EOM pair, the medial and lateral rectus muscles, are quantitatively complex. The interactions of the vertical rectus and oblique EOMs, which also geometrically vary with the orientation of the eye in the orbit, are even more complex. Intuition is an unreliable guide in attempting to understand the cooperative action of the EOMs, and the response to EOM surgery can be misleading. Much of the older literature and even some modern textbooks describe the specific actions of EOMs based on the simplistic assumption that the EOMs pull along straight-line paths from their origins in the annulus of Zinn to their insertions on the sclera. Such assumptions, neglecting the pivotal role of the pulleys as functional origins of the EOMs, lead to serious conceptual errors about nearly all EOM actions and have been a source of much confusion. The varying actions of the EOMs can best be understood using a mathematical model of the muscular and connective tissue forces acting to rotate the eye. As is true of any physical system in static equilibrium, the eye will rotate until the sum of all forces acting on it is exactly zero. It is necessary only to determine the sources of force acting on the eye and the direction of action of each of these forces, and to solve the equations numerically to find eye position. However, the geometry and actions of the EOMs, pulleys, and passive connective tissues are so complex that implementation of such a model requires a digital computer. The Orbit model is such a computational implementation of ocular static mechanics that incorporates realistic geometric anatomy, including pulleys.37 Orbit is available to ocular motor physiologists and strabismus surgeons and provides as its output tabular as well as graphical renderings (such as Fig. 6) of the mechanical state and morphology of the globe and EOMs. The actions of the EOMs generally depend on the starting eye orientation, so broad statements about individual EOM actions represent generalizations to be used cautiously and to which exceptions must be recognized. Intuitively attractive tests of EOM action, such as inferences derived from eye position after the disinsertion or inactivation of an individual EOM, can be confounded by secondary effects of the remaining EOMs on the starting eye orientation directly resulting from the surgical manipulation. The best way to account for EOM action is to use a computational model to determine the components of each EOM's tension acting in each of the three degrees of rotational freedom of the eye. For example, the forces in grams exerted horizontally, vertically, and torsionally by the striated EOMs as computed by the Orbit model are summarized in Table 1 for primary and secondary gaze positions, and in Table 2 for secondary and tertiary gaze positions. The following paragraph illustrates only a small part of the obvious mechanical complexity of the EOMs. The following are generalizations derived from the Orbit model, assuming normal pulley properties for the EOMs. All of these generalizations could change markedly with, for example, the altered pulley locations that can be the cause of some cases of strabismus.38
TABLE 1. Components of Forces in the Striated Extraocular Muscles in Primary
and Secondary Gaze Positions
*Rotations into abduction, elevation, and extorsion are taken as positive. Boldface indicates the maximum force component generated by any EOM in the specified gaze position. LR, lateral rectus; MR, medial rectus; SR, superior rectus; IR, inferior rectus; SO, superior oblique; IO, inferior oblique.
TABLE 2. Components of Forces in the Striated Extraocular Muscles in Primary
and Tertiary Gaze Positions
*Rotations into abduction, elevation, and extorsion are taken as positive. <sd>Boldface indicates the maximum force components of the two EOMs contributing most to the specified gaze position. LR, lateral rectus; MR, medial rectus; SR, superior rectus; IR, inferior rectus; SO, superior oblique; IO, inferior oblique.
The actions of the rectus EOMs are best understood by considering the components of force they generate in primary and secondary gaze positions with the head upright and stationary (see Table 1). The action of the MR is almost purely that of adduction, although the MR has a small torsional action in vertical gaze. Contrary to some recent assertions, the normal MR has minimal vertical action in any secondary gaze position, although surgical transposition of its insertion can give it one. The LR has an action almost purely of abduction, although the LR has a small torsional action in vertical gaze. Again contrary to some recent assertions, the normal LR has minimal vertical action in any secondary gaze position, although surgical transposition of its insertion can give it one. The remaining four EOMs are considered cyclovertical, because they have substantial combined torsional and vertical actions. The actions of the cyclovertical EOMs require consideration of force components generated in both the secondary and tertiary gaze positions (see Table 2). Although the SR is the most powerful elevator, it is also an intorter, most prominently in adducted elevation, where the SR's intorting force is 80% of total force. The SR has a moderate abducting effect, greatest in abducted elevation. The IR is the most powerful depressor, and it is also an extorter, most prominently in adducted depression, where the IR's extorting force is 77% of total force. The IR has a moderate abducting effect, greatest in abducted depression. The oblique EOMs have roughly equal torsional and vertical actions in secondary gaze positions. It can be generally stated that the SO has its greatest vertical action in depression. However, the greatest depressing action of the SO is in adducted depression, and the greatest intorting action in abducted depression. The maximum total tension of the SO is exerted in adducted depression, the position in which it is tested clinically. The SO has minimal horizontal action in primary position; it is a weak abductor in elevation and abducted depression and a moderate adductor in adducted depression. The esotropia increasing in depression that is observed in association with SO palsy is not due to any strong abducting action of the SO, but instead to altered horizontal actions of the remaining EOMs in a setting where the SO palsy has directly produced an excyclotropia. The IO has both its greatest vertical and torsional actions in elevated adduction. Maximum tension in the IO is developed in elevated adduction, the position in which the IO is tested clinically. The IO has minimal horizontal action in primary position; it is a weak abductor in depression and in abducted elevation but is a moderate adductor in adducted elevation. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ACKNOWLEDGMENT |
| Supported by U.S. Public Health Service, National Eye Institute: grant EY-08313 and core grant EY-00331. J. Demer received a Research to Prevent Blindness Lew R. Wasserman merit award and is David and Laraine Gerber Professor of Ophthalmology. |