|
|
| Chapter 1 Anatomy and Cell Biology of the Human Retina ANN H. MILAM , JULIE E. SMITH and SINOJ K. JOHN Table Of Contents |
The term retina derives from the Latin word rete, meaning “net.” This name was originally used by ancient Greeks, who
observed that the retina had the appearance of a fishing net.1 The retina has two functions. The first, performed by the rod and cone
photoreceptors, is to transduce information from an optical image into
electrical signals. The second, performed by the neural circuits of
the retina, is to process certain features of the visual world from the
photoreceptor signals and relay this information to the brain via the
optic nerve.2 Microscopic features of the retina have been studied since the late 1600s, when
Antony Van Leeuwenhoek described globular cells and blood vessels
in a bovine retina.1 By the late 1800s, Ramón y Cajal and Camillo Golgi had developed
silver staining techniques and elucidated the basic cellular organization
of the retina. In his 1892 monograph, La Rétine des Vertébrés, Cajal2 wrote:
|
| DEVELOPMENT OF THE RETINA | ||
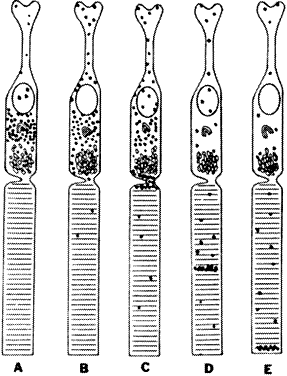
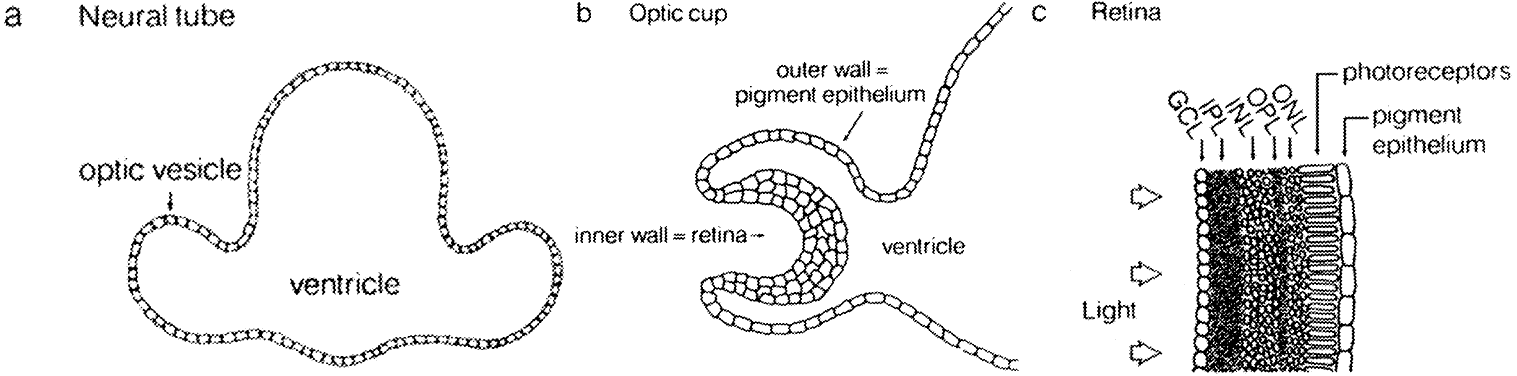
The retina comprises tissues derived from the optic vesicle, an outgrowth
from the diencephalon at approximately 25 days in fetal development (Fig. 1). The optic vesicle invaginates during the fourth fetal week to form a
cup, the inner wall of which differentiates into the multilayered neurosensory
retina (Fig. 2). The outer wall of the cup becomes the pigment epithelium.
|
| RETINAL PIGMENT EPITHELIUM | ||
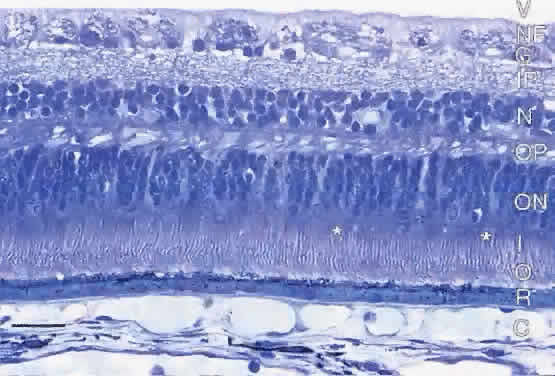
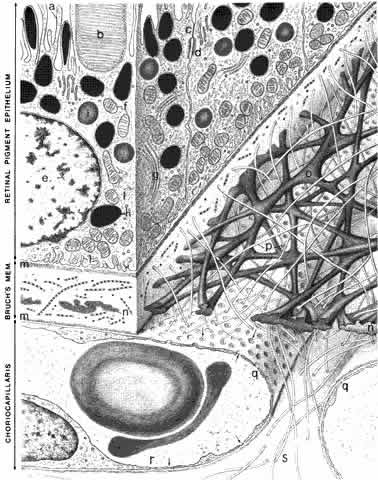
The retinal pigment epithelium (RPE) is a monolayer of hexagonal cells
extending from the margin of the optic disc to the ora serrata, where
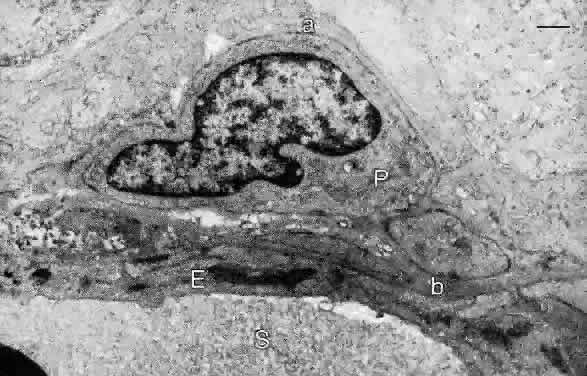
it is continuous with the pigmented epithelium of the pars plana of the ciliary body.3 The RPE cells are cuboidal in cross section and joined near their apical
margins by junctional complexes that include maculae and zonulae adherentes (intermediate spot and belt junctions, respectively), as well as zonulae occludentes (tight belt junctions). In the zonulae occludentes between adjacent RPE
cells, the plasma membranes are fused and are impermeable to water and
ions (Fig. 3). These continuous belts of tight junctions constitute the outer blood—retinal
barrier, which blocks extracellular movement of water, ions, and
macromolecules between the choriocapillaris and the subretinal
space.
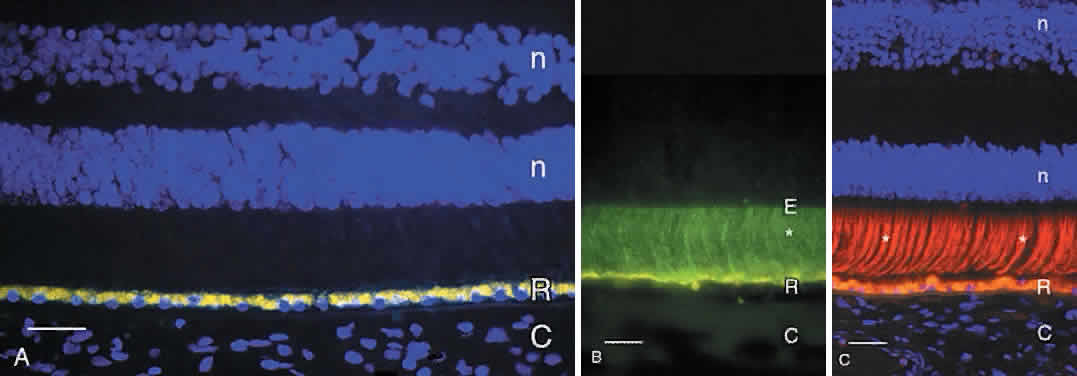
The RPE cells are essential for photoreceptor health and have several other functions, including vitamin A metabolism and regeneration in the visual cycle,4 phagocytosis and degradation of shed outer segment tips, light absorption by melaningranules, heat exchange, secretion of the matrix surrounding the photoreceptor inner and outer segments, and active transport of materials between the choriocapillaris and the subretinal space. The RPE cells are smaller (approximately 10 to 14 μm diameter) and contain more melanin in the macula, and are flatter and wider (up to 60 μmdiameter) with less pigment in the periphery. Photoreceptor density also varies across the retina, but a relatively constant ratio of 45 photoreceptors per RPE cell is found in all retinal regions.5 Each RPE cell is polarized with an apical part adjacent to the subretinal space and a basal portion facing Bruch's membrane. The apical surface has numerous long microvilli that envelop the photoreceptor outer segments and are involved in phagocytosis of shed outer segment tips. The apical portion also contains numerous cytoplasmic filaments, microtubules, and football-shaped melanin granules (see Fig. 3). The melanin is thought to absorb stray light, decreasing light scatter within the eye. The midportion of the RPE cell contains the nucleus, cytoplasmic organelles for protein synthesis (ribosomes, rough endoplasmic reticulum, and Golgi apparatus), smooth endoplasmic reticulum, phagosomes containing outer segment tips undergoing digestion by lysosomal enzymes, and lipofuscin granules. The lipofuscin granules, which increase in number with normal aging, represent residual outer segment lipids incompletely digested by lysosomal activity. The lipofuscin granules are autofluorescent (Fig. 4A) and are relatively sparse in the RPE of children. RPE cells in older eyes are engorged with lipofuscin granules, which also contain incompletely digested melanin granules. The highest density of RPE lipofuscin is found in the macula; this high concentration of lipofuscin granules may compromise RPE functions and has been suggested to cause macular degeneration.6
The lateral and basal surfaces of the RPE cells are highly specialized, containing a variety of different receptors and ion channels. The RPE basal surfaces are infolded, increasing the area of the RPE surface membrane. The basal RPE surface rests on a basal lamina that forms the innermost layer of Bruch's membrane. The other layers of Bruch's membrane are the inner collagenous, elastin, outer collagenous, and the basal lamina of the choriocapillaris (see Fig. 3). The RPE cells have electrogenic sodium-potassium pumps in the apical membrane and chloride-bicarbonate exchange transporters in the basal membrane. The net effect of the pumps, transport systems, and passive ion channels is movement of water across the RPE in a retina-to-choroid direction, promoting adhesion of the neurosensory retina to the RPE.5 |
| INTERPHOTORECEPTOR MATRIX |
| The RPE secretes some of the molecules that fill the subretinal (interphotoreceptor) space. This space, the remnant of the ventricular lumen present during fetal development, is delineated by the apical surfaces of the photoreceptor, Müller, and RPE cells. The outer margin of the space is established by the zonulae occludentes (tight junctions) between the RPE cells, and the inner margin is defined by the external limiting membrane, formed by intermediate junctions (zonulae adherentes) between photoreceptor inner segments and Müller cell apical processes.7 This space is filled with the interphotoreceptor matrix, a heterogeneous collection of proteins/glycoproteins, enzymes, and glycosaminoglycans/proteoglycans that surround the rod and cone inner and outer segments. Notable components of the interphotoreceptor matrix are interphotoreceptorretinoid binding protein (see Fig. 4B), though to function in transport of vitamin A between photoreceptors and the RPE,4 and the matrix sheaths that surround individual rod and cone outer segments. The matrix sheaths of rods and cones differ in composition: those of the rods are stained by certain plant lectins, notably wheat germ agglutinin, while the cone sheaths are positive with other lectins, including peanut agglutinin (see Fig. 4C). The sheaths of the specialized foveal cones are labeled with wheat germ agglutinin and elderberry bark lectins. These matrix sheaths are thought to isolate individual photoreceptor outer segments, maintain outer segment orientation, and contribute to attachment of the photoreceptors to the apical surfaces of the RPE cells.8 |
| PHOTORECEPTORS | |||||||
Human retinas, like those of other vertebrates, contain two types of photoreceptor
cells, rods and cones. Rod cells mediate dim light vision, while
cones function in bright light and are responsible for color vision. The
names of these cells are derived from the shapes of their outer
segments, which contain the visual pigments. Additional differences
are present in the size and shape of the rod and cone inner segments, the
location of the cell bodies, and the size and location of their
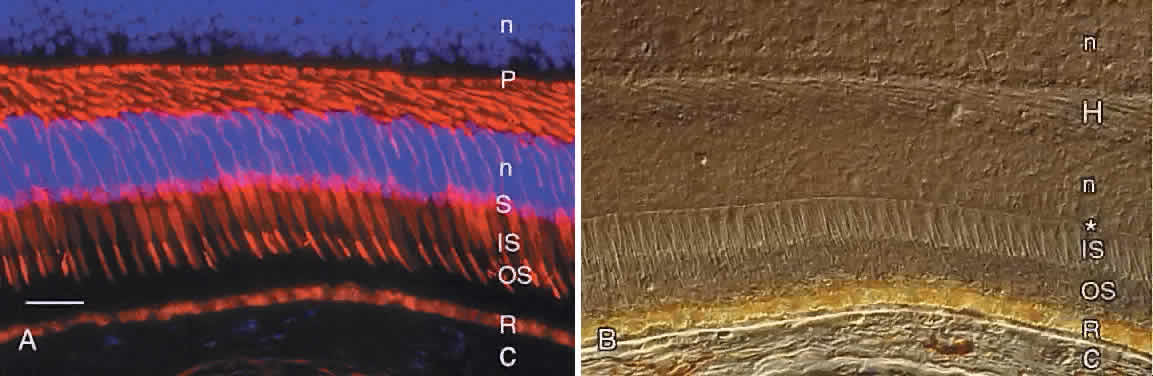
synapses. These features of human cones are revealed by immunocytochemistry
using an antibody against the cone molecule, 7G6 (Fig. 5A). Examination of the same microscopic field by Nomarski differential interference
contrast microscopy reveals other features of the outer retina, including
the external limiting membrane, outer nuclear layer, and
layer of photoreceptor axons and synapses (see Fig. 5B).
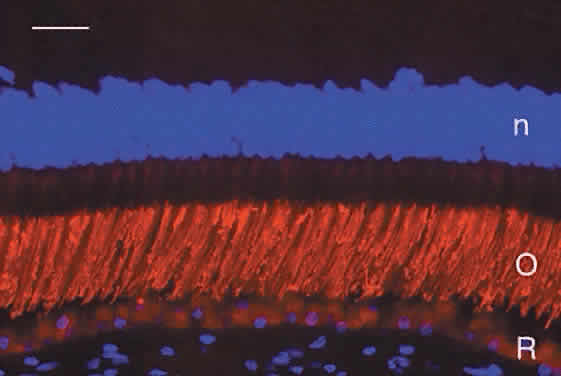
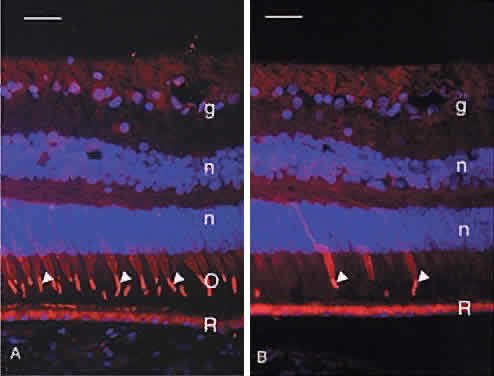
Humans have a single type of rod with a long, thin outer segment containing the visual pigment, rhodopsin, which absorbs maximally at 500 nm. The cone outer segments are shorter and wider and somewhat conical. Each of the three cone types contains a different cone opsin that absorbs maximally in the red (564 nm; L cones), green (533 nm; M cones), or blue (437 nm; S cones) region of the spectrum. The visual pigments can be demonstrated by immunocytochemistry using antibodies specific for the different proteins. Rod outer segments are strongly labeled with anti-rhodopsin (Fig. 6). The composition of the red cone pigment is so similar to that of the green cone pigment that they cannot be differentiated by immunocytochemistry, but the majority of cone outer segments are labeled with an antibody that recognizes both red- and green-sensitive cones (Fig. 7A). A minority of the cones are blue-sensitive (see Fig. 7B).
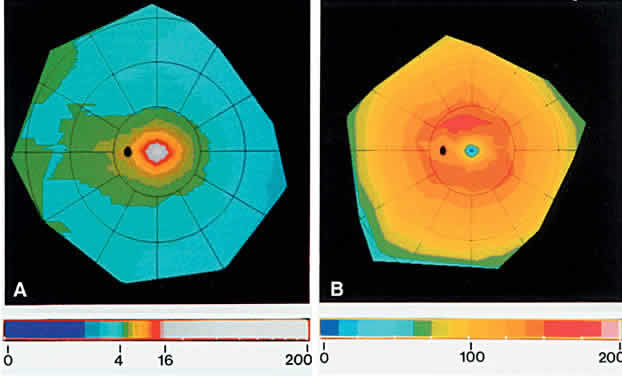
The normal human retina contains approximately 4.6 million cones, with a peak foveal cone density of approximately 199,000 cones/mm2 (Fig. 8). There are approximately 92 million rods, with highest rod density (more than 150,000 rods/mm2) in a ring at the eccentricity of the optic disc.9 Blue-sensitive cones are absent in the foveal center, most numerous (over 2000 cells/mm2) in a ring at 0.1 to 0.3 mm from the foveal center, and constitute approximately 7% of the cones in the periphery.10
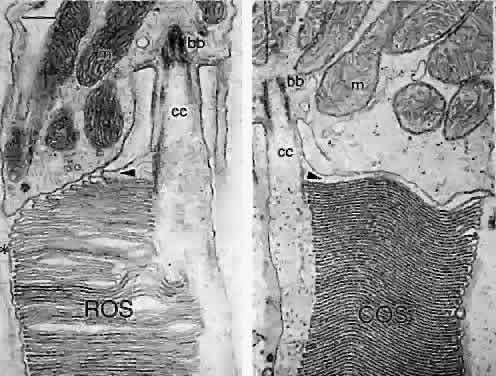
The photoreceptor inner segments contain a cluster of mitochondria forming the ellipsoid, and organelles for protein synthesis (ribosomes, rough endoplasmic reticulum, and Golgi apparatus) in the myoid between the ellipsoid and cell nucleus. The rod and cone outer segments are formed by stacks of membranes containing the visual pigments, rhodopsin and the cone opsins, respectively. In rods, the membranes pinch off at the base of the outer segment to form multiple discs resembling a stack of coins, surrounded by the plasma membrane (Fig. 9). There is ongoing addition of newly synthesized rhodopsin into these membrane discs as they form at the base of the rod outer segment. The newly synthesized protein can be traced by light microscopic radioautography after pulse labeling an animal with radioactive amino acids that are incorporated into rhodopsin being synthesized in the inner segment. The radioactive rhodopsin is then inserted into newly forming basal discs and appears as a dark band across the outer segment in radioautographs. These labeled discs are displaced outward by continuous addition of new, unlabeled discs at the outer segment base (Fig. 10). The process of disc addition is balanced by periodic shedding of the outer segment tip, which is phagocytosed and degraded by RPE lysosomal activity. In primates, complete renewal of a rod outer segment occurs in a about 2 weeks. In cones, the outer segment membranes remain continuous with the surface membrane (see Fig. 9), such that newly synthesized photopigment is inserted diffusely into the outer segment, rather than as a band as found in rods. The time course of cone outer segment renewal is not known.
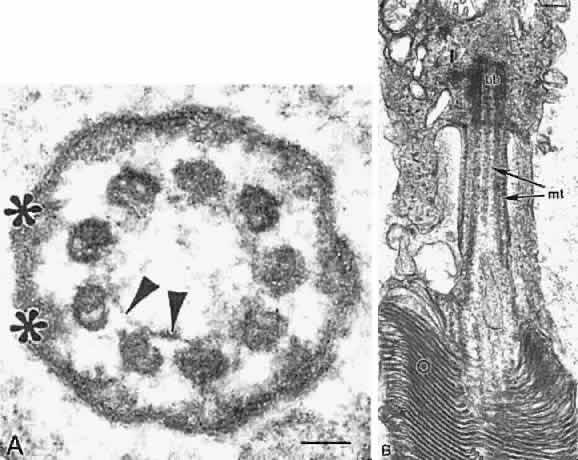
The photoreceptor outer and inner segmentsare joined by a thin stalk, the connecting cilium(Fig. 11). This cilium originates in a basal body (centriole) in the inner segment and as viewed in cross section by electron microscopy contains a ring of nine microtubule doublets typical of a sensory cilium. A central pair of microtubules, characteristic of motile cilia, is absent in the photoreceptor connecting cilium. The cilium is the only connection between the outer and inner segments and is thought to function as a conduit for transport of newly synthesized proteins such as visual pigments from the inner to the outer segment.
|
| EXTERNAL LIMITING MEMBRANE | ||
As described above, a thin membrane is apparent at the level of the photoreceptor
inner segments (see Fig. 5B). This so-called membrane is actually a series of intermediate junctions (zonulae
adherentes) between the rod and cone inner segments and the
apical processes of the Müller cells (Figs. 12 and 13). These junctions constitute the inner border of the subretinal space
and are a barrier to diffusion of large molecules into and out of this
space.7
|
| OUTER NUCLEAR LAYER |
| The outer nuclear layer contains the cell bodies and nuclei of the rods and cones (see Fig. 2). In the parafovea, the outer nuclear layer contains 8 to 10 rows of nuclei that belong mainly to cones. In the remainder of the retina, the outer nuclear layer contains five rows of nuclei, with a single, outermost row of cone nuclei and four rows of rod nuclei internal to them.11 The rod nuclei are smaller and contain denser chromatin than those of the cones. |
| OUTER PLEXIFORM LAYER | |
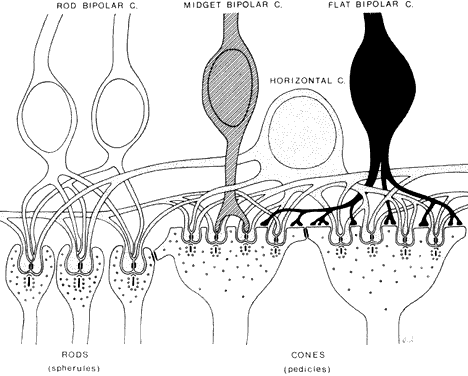
The outer plexiform layer contains the synapses of the rod and cone photoreceptors
onto dendrites of the horizontal and bipolar cells (Fig. 14). The rod synapses are oval and termed spherules. The cone terminals are broader and referred to as pedicles. The cone pedicles form a row internal (vitread) to the rod spherules. The
outer plexiform layer is thickest in the macula, containing axons
of the rods and cones that course radially from the fovea, constituting
the fiber layer of Henle. Both rod and cone synapses contain synaptic
ribbons and vesicles. The rod and cone terminals are organized as invaginating
synapses, termed triads, each containing a single presynaptic ribbon, two lateral processes from
horizontal cells, and a single central process from a bipolar cell. Each
rod spherule contains a single triad; each cone pedicle contains
several triads. Cone pedicles also have small synapses on their surfaces
called flat synapses. These originate from “off” cone bipolar cells and are not
associated with synaptic ribbons. Small lateral gap junctions are also
found between rod and cone terminals, but these are not associated with
synaptic vesicles.
|
| INNER NUCLEAR LAYER |
| The cell bodies of five cell types are found in the inner nuclear layer: horizontal
cells, bipolar cells, interplexiform cells, Müller cells, and
amacrine cells. Each cell type forms a regular mosaic across
the retina, and these cells can be identified anatomically by Golgi staining
and immunocytochemistry with cell-specific antibody markers. The horizontal cells form the outermost row in the inner nuclear layer and their dendrites innervate the photoreceptor axon terminals, forming the lateral elements in the synaptic triads. Three different types of horizontal cells (H1, H2, and H3) are found in human retina, and cells of a given type are electrically coupled by gap junctions to form a widespread syncytium. The large-bodied H1 cells receive input from L and M cones but have little or no input from S cones. Each H1 cell has a long axon with a terminal arbor connected to rod spherules.12 The H2 cells have smaller cell bodies and receive input from all three cone types. Each H2 cell has an axon that contacts mainly S cones. The H3 cell has been reported only in human retina.13 The H3 cell resembles the H1 cell but its dendritic field is larger and often asymmetric. The H3 cells receive input mainly from L and M cones, avoiding S cones, and some H3 cells have axons that descend to terminate in the outer strata of the inner plexiform layer. The horizontal cells are thought to contribute to generation of receptive fields and color opponent properties of bipolar and ganglion cells.14 The bipolar cells receive input from photoreceptors and their axons terminate as ribbon-containing synapses onto amacrine and ganglion cells in the inner plexiform layer. There is a single type of rod bipolar cell, an “on” cell with axon terminals in the innermost stratum of the inner plexiform layer. Each rod bipolar receives input from invaginating synapses in 15 to 20 rod spherules in central retinaand 40 to 50 rod spherules in the periphery.15 There are two main cone bipolar cell types: the “on” cells make invaginating synapses on cone pedicles and have axon terminals in the inner strata of the inner plexiform layer; the “off” cone bipolars make flat synapses onto cone pedicles, and their axons terminate in the outer strata of the inner plexiform layer.16 The cone bipolars are also subdivided as diffuse bipolars that connect to multiple cones versus midget bipolars, each contacting a single cone pedicle. Six diffuse cone bipolar types receive L- and M-cone input, and each foveal cone synapses onto one “on” and one “off” midget bipolar cell. Finally, the “on” blue cone bipolar cell receives input exclusively from S cones.14,17 The interplexiform cells have cell bodies among the amacrine cells and processes that receive input from amacrine cells and make synapses on bipolar and amacrine cell bodies in the inner nuclear layer. The interplexiform cells are unique in being centrifugal, sending processes into the outer plexiform layer to form synapses onto cone pedicles and dendrites of rod and cone bipolar cells.18 The Müller cells are giant glial cells whose processes span the thickness of the retina; these cells are discussed below in the section on glia. The amacrine cells form the innermost row of cells in the inner nuclear layer. There are multiple types (up to 40) of amacrine cells, each with differing morphology and neurotransmitter content.16,19,20 Each amacrine cell type forms a regular mosaic that tiles the retina. The amacrine cells are laterally connected and receive input from bipolar and other amacrine cells in the inner plexiform layer, where they synapse onto dendrites of bipolar, other amacrine, and ganglion cells. |
| INNER PLEXIFORM LAYER | |
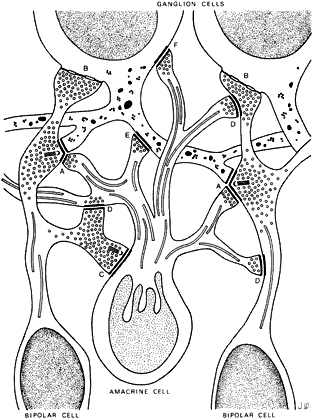
The inner plexiform layer contains distinct strata of synapses from cells
in the inner nuclear layer (Fig. 15). These strata comprise precisely ordered processes and synapses of the
different neuron types. The outer part (sublamina a) contains strata
S1 and S2; the inner part (sublamina b) contains strata S3, S4, and S5. Ganglion
cells that respond to a spot of light when it is turned on ( “on” center cells) have their dendrites in the inner strata, S3, S4, and S5; ganglion cells
that respond to a spot of light when it is turned off ( “off” center cells) have their processes in the outer strata, S1 and S2.21 By virtue of the level of their axon terminals in the inner plexiform
layer, specific bipolar cell types synapse with ganglion cells and amacrine
cells whose dendrites ramify in the same stratum or sublamina. Occasional
cell bodies in the inner plexiform layer are displaced amacrine
or ganglion cells.
|
| ROD AND CONE CIRCUITRY |
| Separate circuits exist for rod and cone signals through the retina. Many
rods converge onto a single rod bipolar cell in the outer plexiform
layer. The axon of a given rod bipolar terminates as a narrow band in
sublamina b of the inner plexiform layer onto dendrites of several amacrine
cell types, including the AII amacrines. The AII amacrine cells
form chemical synapses onto “on” ganglion cells whose dendrites
branch only in sublamina b. The AII amacrines also form electrical
synapses (gap junctions) in sublamina b onto dendrites of “on” diffuse
cone bipolar cells, whose axons synapse on both “on” and “off” ganglion cells. Thus, rod signals reach
both “on” center and “off” center ganglion
cells. Other amacrine cell types are also involved in rod pathways; this
is a complex topic under active investigation. The circuitry for cone signals differs from that for rods in several ways. In the outer plexiform layer, cones synapse onto various cone bipolar cell types rather than onto a single type, as in the rod system. Some cone bipolars synapse in sublamina a of the inner plexiform layer onto “off” ganglion cells, and others synapse in sublamina b onto “on” ganglion cells. Fewer cones converge onto a cone bipolar cell than the numerous rods that input on to a rod bipolar; additionally, relatively few cone bipolar cells converge onto a given ganglion cell. The lowest convergence ratio is found in the midget system in the fovea, where a single L or M cone synapses onto two midget bipolar cells (one “on” and one “off”), which in turn synapse onto two midget ganglion cells (one “on” and one “off”), respectively. This two-to-one relationship between a foveal cone and midget bipolars, with a one-to-one relationship between each midget bipolar and midget ganglion cell, provides the highest level of resolution in the visual pathway. Amacrine cells are also involved in the cone system, modifying the ganglion cell receptive fields and center response. These interactions are complex and not well understood. |
| GANGLION CELL LAYER |
| This layer contains the cell bodies of the ganglion cells, the projection neurons of the retina. There is individual variability in the number of ganglion cells. One study found a total of 0.7 to 1.5 million ganglion cells per retina in six young normal eyes.22 That study also quantified displaced amacrine cells in the ganglion cell layer and found that displaced amacrines accounted for 3% of the total in central retina and nearly 80% in the far periphery. The ganglion cell layer comprises a single row of cells outside the central retina, but within the macula there are up to 10 rows of ganglion cells, with the highest concentration in the parafovea.22 Up to 20 different ganglion cell classes are now recognized, based on Golgi staining, electrophysiology, molecular markers, and central projections.19 The ganglion cell types have characteristic dendritic trees and cell body sizes, and cells of a given class form a regular mosaic across the retina. Each ganglion cell receives synapses onto its dendrites in characteristic inner plexiform strata and sends a single axon into the nerve fiber layer. |
| NERVE FIBER LAYER |
| The nerve fiber layer comprises the ganglion cell axons, which course from
all parts of the retina toward the optic disc to form the optic nerve. Bundles
of the axons are surrounded by Müller and astroglial
processes. The nerve fiber layer is thickest (20 to 30 μm) around
the optic disc, and it thins more peripherally. Axons from nasally located
ganglion cells project directly to the nasal side of the optic disc. Axons
from ganglion cells temporal to the fovea form arcuate fibers
that course above and below the macula and reach the optic disc at its
upper and lower edges. Axons from ganglion cells between the disc and
macula course directly to the disc as the papillomacular bundle. Axons of the nerve fiber layer are unmyelinated and are 0.6 to 2.0 μm in diameter. The cytoplasm of the axons contains microtubules, microfilaments, mitochondria, and vesicles undergoing axonal transport to and from the cell body and central synapses. After passing through the lamina cribrosa of the optic disc, the individual ganglion cell axons become myelinated and the diameter of the optic nerve expands at this point. |
| INTERNAL LIMITING MEMBRANE |
| The inner surface of the retina is covered by a basal lamina derived from the Müller glial cells. It measures 1 to 2 μm in thickness and is positive by periodic acid-Schiff staining. This membrane is closelyassociated with the Müller end feet and merges with collagen fibrils of the vitreous. |
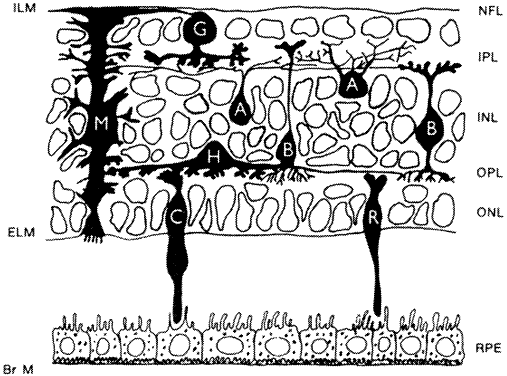
| GLIA MÜLLER CELLS | |||
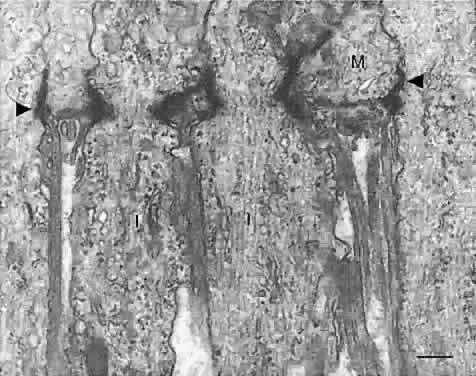
Müller cells are radial glia that extend from the vitreal surface
of the retina to the subretinal space(Fig. 16). The Müller cell body and nucleus are located midway in the inner
nuclear layer. An apical cell process extends to the external limiting
membrane, where it ends as microvilli (the fiber basket of Schultze) projecting
into the subretinal space around the photoreceptor inner
and outer segments. The external limiting membrane comprises bands of
intermediate junctions between the Müller apical processes and the
photoreceptor inner segments (see Figs. 12 and 13). A basal Müller process extends from the cell body to the vitreal
border, where it terminates as end feet on its basal lamina, the inner
limiting membrane. Müller cells send out fine cytoplasmic processes
into the retinal parenchyma, where they surround all neurons and
their processes, particularly in the two plexiform layers. The only sites
where retinal neurons directly contact each other, without intervening
Müller cell processes, are at their synapses. In the nerve
fiber layer, Müller processes surround bundles of ganglion cell axons. Müller
cell end feet also terminate against the basal laminae
of blood vessels in the inner retina.
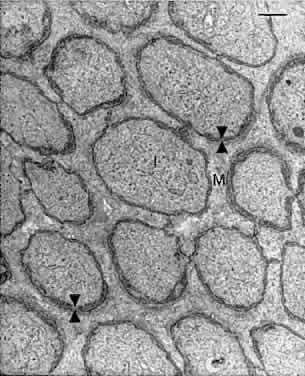
For many years, the Müller cells were thought to function mainly for structural and metabolic support of the retinal neurons. Recent studies have disclosed additional Müller cell functions, including regulation of extracellular potassium levels, uptake and degradation of neurotransmitters, and regulation of CO2 and pH levels in the retina.23 Müller cells also contain several binding proteins for vitamin A, suggesting that they may play a role in the visual cycle.4,24 ASTROCYTES As in brain, the astrocytic glia are stellate cells with oval nuclei. Their cell bodies are located mainly in the nerve fiber layer, with a few in the ganglion cell layer. The astrocytes can be demonstrated by immunocytochemistry with an antibody against glial fibrillary acidic protein (Fig. 17). The astrocytes are prominent in the prelaminar portion of the optic disc, and wide astrocytic foot processes surround the walls of the retinal blood vessels.
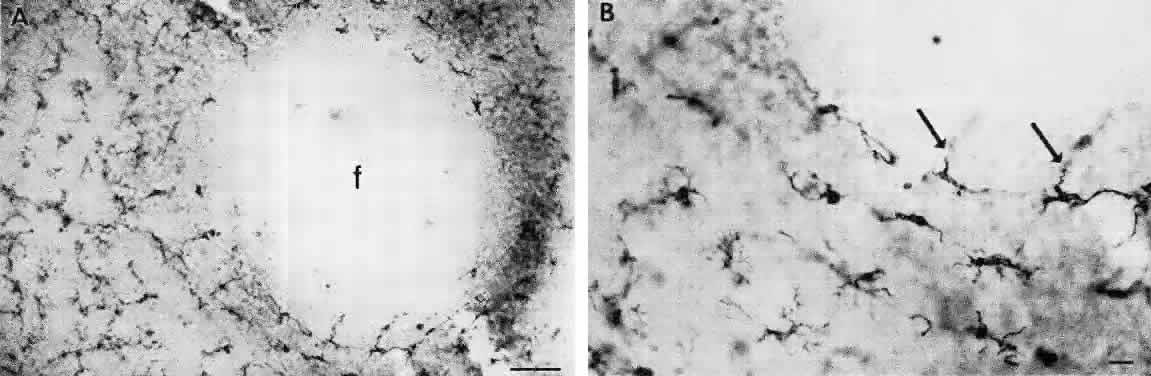
MICROGLIA These stellate cells form regular mosaics in the interfaces between the nerve fiber and ganglion cell layers and the inner nuclear and outer plexiform layers.25 The microglia are tissue macrophages derived from the bone marrow. They are closely associated with the retinal blood vessels and are absent from the avascular foveal pit (Fig. 18). The microglia can be demonstrated with CD45 and HLA-DR leukocyte markers.26 The microglia become activated in damaged or degenerating retinas, where they function as scavengers of debris and may function to present foreign antigens to T lymphocytes.27
|
| VASCULAR ELEMENTS | |
| The retina has a dual blood supply, and there are two blood—retinal
barriers. The RPE and photoreceptor layers are nourished by the choriocapillaris, whose
endothelial cells are fenestrated and leak serum
proteins that diffuse along the lateral walls of the RPE cells. Tight
junctions between the RPE cells block diffusion of serum components into
the subretinal space and constitute the outer blood—retinal barrier. In
contrast, endothelial cells of the central retinal circulation
lack fenestrations and are joined by zonulae occludentes, forming
the inner blood—retinal barrier. The inner nuclear and ganglion cell layers are supplied by branches of the central retinal artery and vein, which divide to supply the four quadrants of the retina. Within the retina, the main arterial and venous branches lie in the nerve fiber layer and give off arterioles and venules within this layer. The capillaries arising from the arterioles in the nerve fiber layer pass directly into the retina, often at right angles to the vessel of origin. They connect with venous capillaries at all levels in the inner retina. The arterial branches have smooth muscle walls and an endothelium but lack an internal elastic lamina. The branches of the central retinal vein have thin walls of smooth muscle and an endothelium, and the walls of the venules and capillaries contain mural cells (pericytes) enclosed within a basal lamina continuous with that of the endothelial cells (Fig. 19). The endothelial nuclei are oriented along the capillary axis and are long and pale; the mural cell nuclei are small and dense.
The peripapillary retina has a distinct group of capillaries forming a network in the nerve fiber layer. These capillaries course along the vessels and nerve bundles, following the arcuate distribution of ganglion cell axons above and below the macula. In the macula, capillaries arise from arterioles off the superior and inferior temporal arteries. These capillaries divide and subdivide at all levels of the inner retina in the macula, except for the fovea, where they are absent in an area 0.25 to 0.6 mm in diameter. As described below, the foveal pit contains only photoreceptors and RPE cells, which are nourished by the underlying choriocapillaris. Lateral displacement of the ganglion cell and inner nuclear layers to the parafovea obviates the need for central retinal vessels within the fovea. This specialization contributes to increased visual acuity in the fovea because blood cells do not circulate between the incoming light and the photoreceptors. |
| REGIONAL ANATOMIC VARIATIONS | ||||
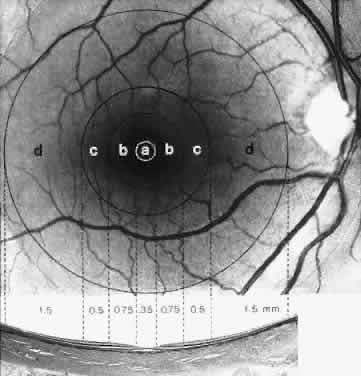
MACULA The meaning of the terms macula lutea (yellow spot), fovea centralis (fovea), and foveola varies, depending on whether they are used in a clinical or histologic context. Clinically, the macula is identified as the yellow, capillary-free zone temporal and slightly below the center of the optic nerve head. The yellow color of the macula is due to its content of two xanthophyll pigments, lutein and zeaxanthin, derived from carotenoids in the diet.28 These pig-ments are antioxidants and are thought to protect the fragile photoreceptor outer segments from photo-oxidation and to absorb phototoxic short-wavelength light. By absorbing blue light, the macular pigments may also decrease chromatic aberration in the macula. In the center of the yellow region, an annular light reflex defines the fovea. The tiny reflex in the center of the foveal reflex identifies the foveola. Histologically the macula (diameter 5.5 mm) is defined by several features and is subdivided into the fovea, foveola, parafovea, and perifovea (Fig. 20).
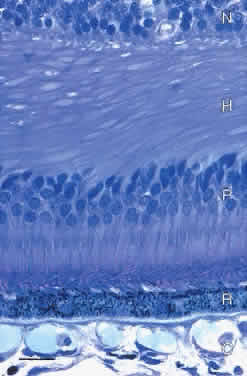
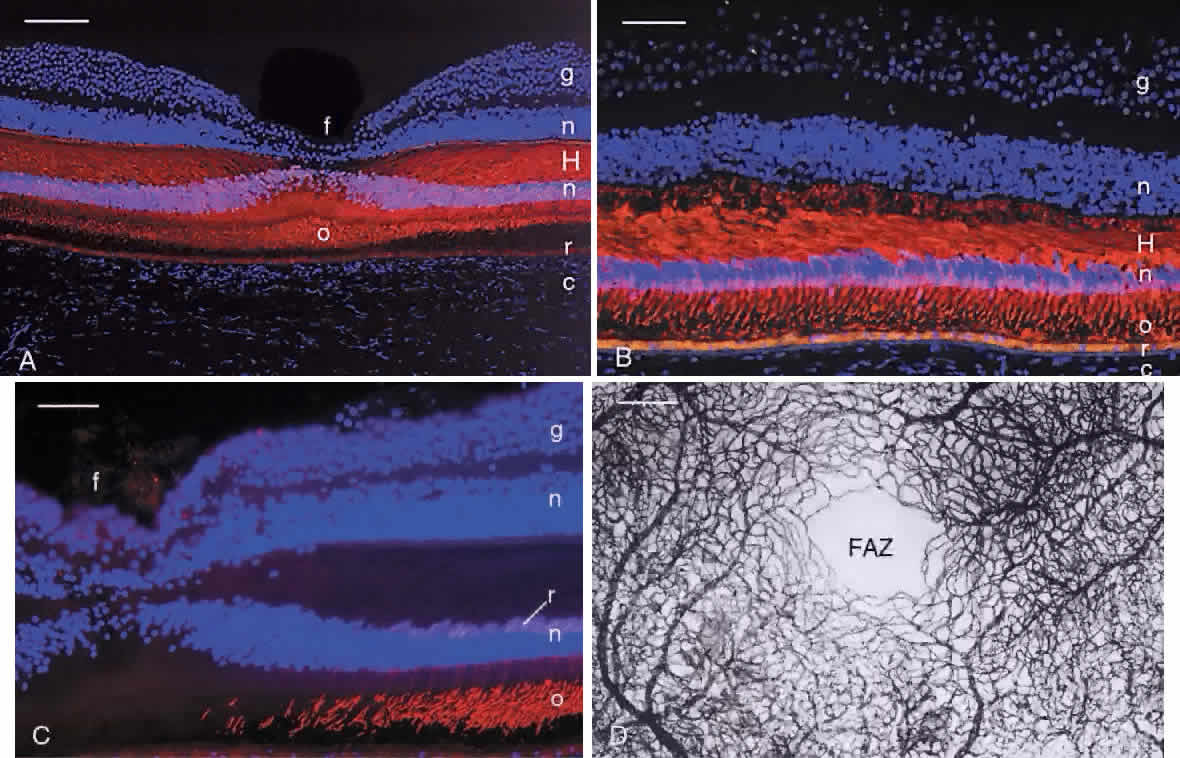
FOVEA The central fovea (fovea centralis) is a small pit in the vitreal surface of the retina (Fig. 21). Its center is located 4.0 mm temporal and 0.8 mm inferior to the center of the optic disc. The fovea is approximately 1.5 mm in diameter and the depth of the fovea is variable but on the order of 0.25 mm. In the center of the fovea the retina is reduced in thickness to approximately 0.13 mm. The inner nuclear layer is reduced to two rows of cells at the edge of the fovea and is absent in the foveal pit. The inner plexiform, ganglion cell, and nerve fiber layers are also absent from the pit.
The photoreceptor layer in the center of the foveal pit (the foveola, approximately 200 μm in diameter) contains only cones (see Fig. 21). These cones are specialized for high-acuity vision. Their outer segments are approximately 2 μm wide and 45 μm long and tightly packed for increased resolution. The inner nuclear layer cells are displaced laterally so that photoreceptor axons in the outer plexiform layer course radially to synapse onto horizontal and bipolar cells. The thick layer of radial axons is termed the fiber layer of Henle (Figs. 22 and 23). Rods with long, thin outer segments are present in the slope of the wall of the fovea (see Fig. 23C). The central rod-free area is 350 to 600 μm in diameter.1,9
Capillaries are present in the inner retinal layers to the edge of the fovea. The capillary-free zone (foveal avascular zone) is 0.25 to 0.6 mm in diameter. The foveal avascular zone is an important landmark in fluorescein angiography to localize the center of the fovea. The parafovea is approximately 0.5 mm wide, including the foveal region, and measures about 2.5 mm in diameter (see Fig. 20). This is the thickest part of the retina due to lateral displacement into this region of ganglion cells and inner nuclear layer neurons during development of the fovea. The inner nuclear layer contains up to 12 rows of cells and the ganglion cell layer contains up to 10 rows of cells. The fiber layer of Henle is also quite thick in this region and comprises the axons of both rods and cones. The perifovea measures 1.5 mm in width, and the entire macular region measures 5.5 mm in horizontal diameter. This region ends where ganglion cells are reduced to a single row, as found elsewhere in the retina. ORA SERRATA The boundary between the retina and the ciliary body is termed the ora serrata because the retinal edge forms dentate processes that encroach on the pars plana of the ciliary body. The peripheral retina becomes attenuated as it approaches the ora serrata. The photoreceptor layer is reduced to two or three rows of cells, and the inner and outer nuclear layers become gradually fused. Rods are absent about 1 mm posterior to the ora, and the remaining photoreceptors are stunted cones lacking outer segments.29 With increasing age, cystoid spaces form in the plexiform layers. These spaces are surrounded by Müller cells and filled with acid mucopolysaccharide. Peripheral retinal degeneration may occur because the ora serrata is in a watershed zone between the anterior and posterior vascular systems. The transition from the neurosensory retina to the ciliary body epithelium is rather abrupt. The inner limiting membrane in the far periphery is thick and provides a firm attachment for the collagen fibers of the vitreous base. |